
記住我




The involvement of mRNAs and lncRNAs in a variety of biological procedures has been extensively documented in cancer development via rapid advances in sequencing techniques. Consequently, it is imperative to discern the differentially expressed lncRNA or mRNA profiles, and the subsequent lncRNA-mRNA regulatory axis is essential in ESCC progression, which accelerates the disclosure of novel molecular targets and prognostic indicators [22,23,24]. Although numerous lncRNAs play vital roles in the development and metastasis of ESCC, there is a lack of knowledge regarding novel lncRNAs or their involvement in miRNA or mRNA regulation [25]. In the current study, the interaction between the co-expression of markers was explored to clarify the lncRNA-mRNA axis that is associated with ESCC development [26].
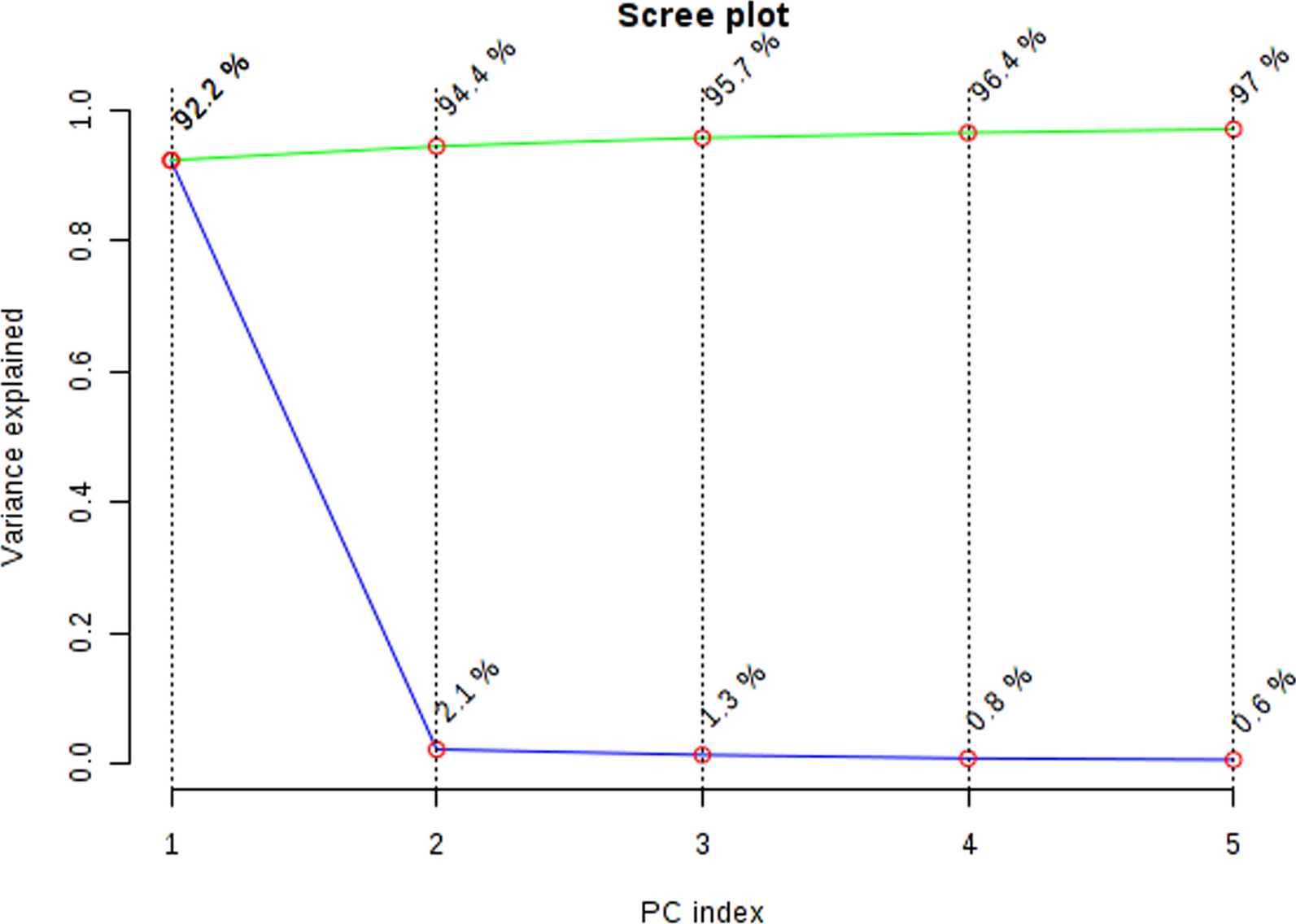
Here, the first results of the expression level indicated the upregulation of AGAP2-AS1, DLEU2, CD44S, TWIST1, and MMP13 and the downregulation of HMBOX1_1 and UGDH-AS1 in ESCC tissue patients and their remarkable association with the clinicopathological characteristics of patients. Given these correlations, we postulate that these lncRNAs and EMT-related coding mRNAs may be involved in tumor formation, EMT procedure, metastasis, and tumor-developing phenotype in ESCC patients. Moreover, we revealed a considerable correlation between the concomitant expression of AGAP2-AS1, DLEU2, CD44S, TWIST1, MMP13, HMBOX1_1, and UGDH-AS1 with each other, hypothesizing that the concomitant expression of transcripts can be related to the possible function of all of them. It is postulated within the realm of scientific inquiry that the concurrence of gene expression and lncRNA transcription plausibly indicates a functional association [27]. The possible correlation between AGAP2-AS1, DLEU2, HMBOX1_1, and UGDH-AS1 and EMT-related genes facilitates the regulation of a multitude of tumor signaling pathways, thereby identifying factors that induce EMT and promote tumor metastasis. This, in turn, enables the identification of appropriate treatment options for epithelial-mesenchymal tumors (Fig. 4) [28, 29].
Fig. 4
The probable crosstalk between AGAP2-AS1, DLEU2, HMBOX1_1, and UGDH-AS1, with EMT markers (CD44S, TWIST1, and MMP13) during the EMT process. EMT markers possess the ability to govern the expression of their subsequent genes as well as MMPs [33]. The dysregulation of MMP13, CD44S, and TWIST1 exhibits a significant association among themselves [16]. The expression levels of GAP2-AS1, DLEU2, HMBOX1_1, and UGDH-AS1 are significantly associated with each other and EMT markers
Reducing ESCC-associated mortality necessitates a thorough comprehension of the functional roles of relevant genes in various signaling pathways, which could potentially enable the creation of a gene panel supported through preclinical explorations and clinical validation [5, 30]. Dysregulation of numerous protein-coding genes, signaling pathways, and epigenetic factors can mediate EMT status as a factor in tumor invasion [31]. EMT is induced through a series of TFs (such as SLUG, ZEB1/2, TWIST1/2, SNAIL1/2, and SIP1) that regulate EMT markers (Fibronectin, N-cadherin, E-cadherin, Claudins, Vimentin, and Occludins) as well as matrix metalloproteinases (MMPs) that degrade the extracellular matrix (ECM) and the cell microenvironment [32]. Similar to previous studies, our results confirmed the significant co-expression of CD44S, MMP13, and TWIST1 with patients’ clinicopathological features in ESCC, indicating their role at the beginning of tumor cell invasion. The correlation between the co-expression of CD44S, MMP13, and TWIST1 was significantly related to tumor development, poor prognosis, and decreased patients' overall survival, proposing a potential crosstalk between these genes in the pathogenesis of ESCC [16]. The manifestation of a CSC-like phenotype strongly correlates with the activation of EMT, thereby improving the proliferation, differentiation, and self-renewal capacity of the tumor-initiating cells [33]. Our results are consistent with previous studies that provided evidence for the role of CD44S and TWIST1 in CSC/EMT status [34]. It has been confirmed that the correlation between CD44S (as a CSC marker), TWIST1 (as an EMT signaling-related gene), and MMP13 (as a collagenase) has a significant role in MET, and the CD44S-TWIST1-MMP13 axis is involved in ESCC aggressiveness [16, 19]. On the other hand, considering the established function of lncRNAs in the EMT process during the development of tumors, it is unsurprising that numerous lncRNAs are additionally linked to the control of cancer phenotypes [35]. It is noteworthy that lncRNAs exhibit a more tissue-specific expression pattern compared to protein-coding genes. Consequently, discovering ESCC-related lncRNAs and investigating their clinical significance as well as their biological role may prove to be a more sensitive biomarker [15, 36].
Interestingly, the upregulation of AGAP2-AS1 was negatively related to poor prognostic results in numerous cancer types, including glioma, glioblastoma, pancreatic (PC), gastric cancer (GC), colorectal cancer (CRC), non-small cell lung cancer (NSCLC), papillary thyroid, epithelial ovarian, and hepatocellular carcinoma (HCC) [37]. It has been informed that AGAP2-AS1 silencing promotes cell apoptosis and inhibits cell proliferation, invasion, and metastasis via interaction with EZH2 and LSD1, as well as inhibition of LATS2 and KLF2 expression in NSCLC; consequently, AGAP2-AS1 was recognized as an oncogene [38]. Moreover, AGAP2-AS1 silencing repressed esophageal cancer (EC) development via downregulation of Fos-like antigen-1 (FOSL1) and upregulation of miR-195-5p, suggesting that AGAP2-AS1 can lead to EC progression [39]. Additionally, the upregulation of exosomal AGAP2-AS1 has been indicated to prevent trastuzumab-induced cell cytotoxicity through packaging into exosomes in HER2+ breast cancer cells [40]. Our date along with previous results, showed that the upregulation of AGAP2-AS1 was correlated with tumor development in ESCC. Given the oncogenic role of TWIST1, MMP13, and CD44S and their upregulation in ESCC [16], we have also observed similar outcomes concerning their concomitant expression with AGAP2-AS1. Our findings indicated that the overexpression of AGAP2-AS1 was associated with TWIST1, MMP13, and CD44S overexpression in the primary stages of ESCC development, and tumor cells with poorly and moderately differentiated invaded the adventitia (T1/T2) with lymph node metastasis. The empirical evidence has shown that the TWIST1 upregulation leads to the MMP13 upregulation, ECM remodeling, EMT, and tumor progression [41]. The data gathered in conjunction with previous research elucidates the role of AGAP2-AS1 in EMT and the degradation of ECM components, proposing that AGAP2-AS1 plays a crucial role in the invasion and metastasis of ESCC.
For the first time, our investigation revealed the occurrence of UGDH-AS1 downregulation in ESCC. This dysregulation was found to be linked with tumor invasion, stages of tumor progression, and tumor metastasis. Based on GEO and TCGA database analysis, the downregulation of UGDH-AS1 was correlated with patient overall survival [25]. Our findings demonstrated a noteworthy association between the co-expression of UGDH-AS1 with TWIST1, CD44S, and MMP13 during the initial steps of tumor infiltration and progression. Induction of TWIST1 expression through CD44S leads to the EMT process and can propose an oncogenic role for UGDH-AS1 in ESCC [42]. Therefore, our findings may pave the way for additional research aimed at enhancing our comprehension of the conceivable molecular mechanism of UGDH-AS1 in ESCC tumorigenesis.
Given that DLEU2 is upregulated in HCC, glioma, CRC, cervical, laryngeal squamous cell carcinoma (LSCC), and NSCLC [43,44,45,46,47,48]; we found similar results for this lncRNA in ESCC. The results of our study have demonstrated a notable association between the overexpression of DLEU2 and the upregulation of TWIST1 and CD44S during the advanced stages of ESCC progression. This relationship was particularly evident in tumor cells with a well-differentiated grade, which exhibited invasive behavior into the lamina propria, muscularis propria (T3/T4), and metastasis into the local lymph nodes. It has been reported that DLEU2 predicted poor prognosis, promoted proliferation/migration/invasion, increased apoptosis, and enhanced malignant properties via regulation with EZH2, Bcl-2/Bax axis, miR-30e-5p/E2F7 axis, caspase cascade, miR-30c-5p/SOX9 axis, miR-186-5p/PDK3 axis, DLEU2/miR-455/SMAD2 pathway, miR-30c-5p/PIK3CD/Akt axis, miR-128-3p, and STAT1/DLEU2/miR-23b-3p/NOTCH2/Notch axis in HCC, EC, NSCLC, glioma, PC, LSCC, cervical cancer, GC, and CRC [45,46,47,48,49,50]. Consequently, the upregulation of DLEU2 as an lncRNA with numerous splice variants is associated with poor survival rates in lung, pancreatic, esophageal, cervical, laryngeal, and renal cancers and seems to act as an oncogene in most tissues. The results of previous studies represent evidence for the valuable prognostic and therapeutic role of this lncRNA as a novel biomarker in malignancies [51].
The TCGA database analysis identified HMBOX1_1 (AC108449.2) as an immune-related lncRNA signature and an independent prognostic lncRNA for overall survival rate in kidney renal clear cell carcinoma (KIRC) patients, where its downregulation was negatively related to tumor staging [52, 53]. Moreover, Cox regression analysis exhibited a significant correlation between the upregulation of autophagy-related HMBOX1_1 (AC108449.2) and overall survival among high-risk bladder urothelial carcinoma (BLCA) patients [54]. However, there are no reports on the functional role, dysregulation, clinical importance, and correlation between the HMBOX1_1 (AC108449.2) expression with patients’ clinicopathological parameters in various types of malignancies. Our data showed underexpression of this lncRNA and its co-expression with CD44S and MMP13 markers in advanced steps of tumor development, where data document its correlation with clinicopathological parameters in ESCC for the first time. CSC markers (such as GLI1, CD44, and SALL4) have been found to be expressed in tumor cells during tumor progression. In light of the CD44 stem cell-associated surface marker, it can be posited that it expedites the emergence of the EMT phenotype by means of galvanizing mesenchymal markers and EMT-TFs [55]. Taken together, the downregulation of HMBOX1_1 (AC108449.2) may activate the CD44S-TWIST1-MMP13 axis to promote EMT and invasion in ESCC.
This study is a descriptive study by qRT-PCR that confirms the protein expression levels by immunoblotting and/or IHC. Another limitation of this study is the number of samples, as a large number of tissue samples are required to verify the outcomes. Considering the effect of the CD44-TWIST1-MMP13 axis in the EMT process, evidence to confirm this effect using molecular methods is needed, so molecular mechanistic studies using ESCC cell lines would be desirable. In addition, considering that the strength of metastasis also increases, it is necessary to investigate the migration and invasion experiments.
留言 (0)