
記住我
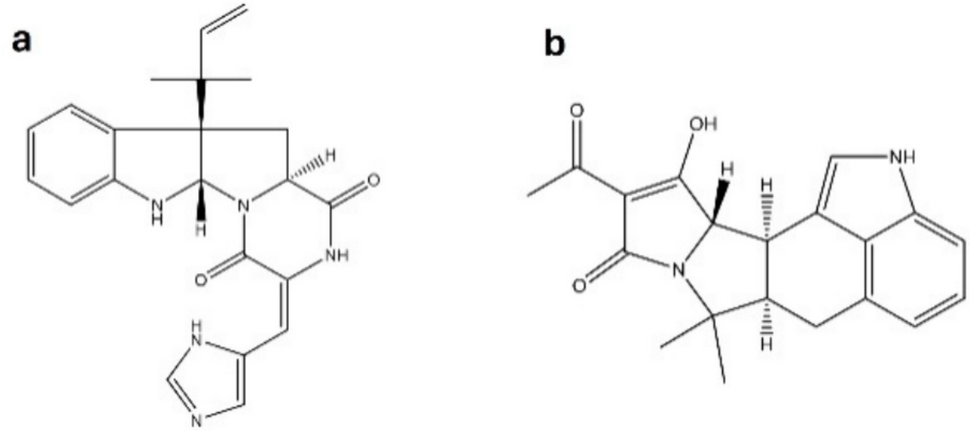




To achieve a fast, high-throughput and cost-effective TTR competitive binding assay, experimental conditions were optimized aiming for an ultimate balance between proper functioning of the assay, physiological relevance, and practical ease. Therefore, first the impact was tested of different incubation temperatures (on ice (4 ºC) and room temperature (22 ºC)) and incubation times (5, 15, 60, and 120 min (at room temperature)) on the obtained Kd value for FITC-T4 and the Ki values of the selected test items. In a next step, different FITC-T4 concentrations (changing the FITC-T4:TTR ratio) were assessed.
The saturation curves of the FITC-T4-TTR complex at different incubation conditions and their corresponding Kd values are shown in Fig. 2 and Table 2, respectively. The Kd value of the FITC-T4-TTR complex at room temperature (143 nM, CI 127–159 nM) did not differ significantly from the Kd value on ice (157 nM, CI 124–190 nM), as the 95% confidence intervals overlap (Table 2). The Kd values of the FITC-T4-TTR complex also did not significantly change with different incubation times (Table 2). To demonstrate that differences in temperature and exposure time do not affect the outcomes for test items, competitive TTR-binding experiments were performed with T4, PFOS, and PFOA at different temperatures and incubation times (Fig. S1 and Table S1). For T4, PFOS, and PFOA, concentration response curves and corresponding Ki value estimations for competitive TTR-binding did not differ significantly between different incubation conditions.
Fig. 2
TTR saturation binding curves obtained with different experimental conditions. Assays were performed with 30 nM TTR at a different incubation temperatures (RT = room temperature) analysed after 120 min and b different time points at room temperature (N = 3, n = 3). Data are presented as the average (± SD) of separate experiments
Table 2 Kd values of the FITC-T4-TTR complex at RT and on ice after 120 min of incubation and at different time points at room temperature (RT). (CI = confidence interval, N = 3, n = 3)The results observed from both the saturation and the competitive binding experiments indicated that incubations on ice (at 4 ºC) can be replaced by incubations at room temperature, since no significant differences in the Kd and Ki values were obtained at two incubation temperatures. Kd and Ki values were not determined at 37 ºC, which may be considered more physiologically relevant. However, incubations at 37 ºC would make the protocol less suited for daily routine measurements as laboratories may lack plate readers with a heating element. Besides, testing at 37 ºC is not expected to provide more relevant data because it was reported that that FITC-T4 displacement from TTR by T4 at 37 ºC did not differ significantly from FITC-T4 displacement at 4 ºC or RT (Hamers et al. 2020). Additionally, the different incubation time points also did not significantly affect the Ki values or the binding potency of the test items. Although shorter incubation time (5 min) worked adequately, for practical reasons an incubation time of 15 min was chosen enabling easy screening of multiple plates by one technician (i.e., leaving sufficient time between different plates to perform the FI measurements).
In a next step, the impact was assessed of different FITC-T4 concentrations (11, 55 and 110 nM) in combination with 30 nM TTR (changing the TTR:FITC-T4 ratio) on competitive binding of T4 and PFOS (Fig. S2). The impact of the FITC-T4 concentration used is limited and using about 11 nM FITC-T4 still resulted in adequate curves. However, although the instrument used is sensitive enough to measure a low concentration of FITC-T4, instruments at other facilities might not be that sensitive. Therefore, in order to develop an assay that can be used at different labs, the concentrations of TTR and the FITC-T4 label were kept at 30 and 110 nM, respectively (i.e., the same as used before (Hamers et al. 2020)).
Eventually the assay turned out to work adequately at room temperature with an incubation time of 15 min and by using final concentrations of 30 nM TTR and 110 nM of the FITC-T4 label. Under these conditions, the Kd value of the FITC-T4-TTR complex is 142 nM (Table 2). The IC50 value of T4 with the FITC-T4-TTR complex is 125 (CI: 105–146) nM and the corresponding Ki value is 41 (CI 34–49) nM (Fig. 4, Table 3). These values are similar to IC50 = 100 nM and Ki = 32 nM, as previously reported (Hamers et al. 2020). These results demonstrate that the optimized protocol can be used for an easy, physiologically relevant, and fast in vitro testing of compounds for their capacity to disrupt T4 binding to TTR.
Table 3 IC20, IC50 and Ki values (nM) of the seven model compounds with TTR (N = 3, n = 3) and TBG (N = 5, n = 2 for T4 and N = 3, n = 3 for other compounds) from the present study and from the literatureDevelopment of a TBG-binding assayBased on the optimized TTR-binding assay, a new TBG-binding assay was developed in the present study using FP as a readout. Initially, FI was used as a readout for the development of the TBG-binding assay to mimic the optimized TTR-binding assay as closely as possible. However, the saturation curve of the FITC-T4-TBG complex (30 nM TBG with different concentrations of FITC-T4) showed large variations for replicates within a single experiment (Fig. S3a). In addition, the signal to noise ratio (FI values of experimental wells divided by that of background wells) was very low and around 1.0 only (Fig. S3b). Upon binding to TBG, the increase in FITC-T4 was insufficient to distinguish TBG-bound from free FITC-T4, which may be explained by the different binding site of TTR and TBG for T4 (Refetoff 2023; Ren and Guo 2012). In order to increase the signal to noise ratio, a higher concentration of TBG (60 nM) was tested in the saturation experiment (Fig. S3b). With 60 nM TBG, the signal to noise ratio slightly increased compared to that with 30 nM TBG, but was considered not high enough to eventually obtain a robust TBG-binding assay using FI as readout. Therefore, FP was tested. It is considered as an alternative readout, as it has been demonstrated before to be a successful technique to study interactions between ligands and proteins (Rossi and Taylor 2011). In contrast to FI, where the emission intensity is measured, with FP the molecular Brownian rotation during the time between excitation and emission of the tracer is measured. FP is often expressed as anisotropy (r), which is defined as:
with \(_\) the fluorescence intensity emission parallel and \(_\) the fluorescence intensity emission perpendicular to the excitation direction. The basic principle of FP is that a small molecule, like unbound FITC-T4, rapidly rotates in solution resulting in a lower anisotropy, while a slower rotation is observed when FITC-T4 is bound to a larger molecule, like TBG in the present study, resulting in a higher anisotropy. When competitors are introduced to the FITC-T4-TBG complex, FITC-T4 is displaced, resulting in a decrease in anisotropy. Because the anisotropy signal is a weighted average of the bound and free FITC-T4 signals, saturation experiments with FP readout usually do not titrate increasing concentrations of ligand against a fixed concentration of protein, to avoid that the free ligand signal exceeds the signal of the bound ligand (Nosjean et al. 2006). Instead, saturation experiments are usually performed by titrating increasing concentrations of protein against a fixed concentration of ligand. Therefore, the saturation experiment for TBG had a different design than that for TTR: a fixed concentration of FITC-T4 and a range of concentrations of TBG were used.
Using 5 nM FITC-T4, the signal-to-noise ratio (FP values of wells to which 300 nM TBG was added divided by that of wells containing no TBG) of the saturation curve increased by as much as sixfold, indicating that for TBG the FP readouts have a wider dynamic range than FI readouts, allowing quantification of the binding capacity of potential competitors. In a preliminary TBG saturation experiment (Fig. 3) and competitive binding experiment (Fig. S4) two temperatures were tested: RT and 37 °C (to mimic the human in vivo situation) using an incubation time of 15 min. In this saturation experiment, the Kd value was determined to be 0.5 (CI 0.1–0.8) nM at RT and 1.3 (CI 0.4–2.2) nM at 37 °C, respectively (N = 2). Higher Kd values were observed with increasing temperature, which may be explained by the temperature-sensitive flexibility of the binding pocket (Qi et al. 2014, 2011). Concentration–response curves of test compounds T4 and triclosan did not significantly differ when tested at RT or 37 °C (Fig. S4, N = 2). Correspondingly, the Ki values at these two temperatures for T4 were estimated to be 2.9 nM (CI 0.2–5.6 nM) and 2.6 nM (CI 1.5–3.7 nM) respectively, and 192 nM (CI 178–206 nM) and 175 nM (CI 161–188 nM) for triclosan. Considering the small absolute difference in the Kd values and the similarity in concentration–response curves at the two different temperatures as well as practical concerns as described before for the TTR-binding assay, RT was also selected as the incubation temperature for the TBG-binding assay.
Fig. 3
TBG saturation binding curves assays obtained with 5 nM FITC-T4 at two different temperatures (RT and 37 °C) (N = 2, n = 2, data are presented as the average (± SD) of separate experiments). The inserted graph shows the same data plotted with a logarithmic scale on the x-axis
Saturation curves of the FITC-T4-TBG complex at RT were performed three times (Fig. S5) resulting in an average Kd value (CI) of the FITC-T4-TBG complex of 1.1 (CI 1.0–1.2) nM. In a next step, the concentration of TBG to be used in the competitive binding assay was selected. For competitive TBG binding studies, a concentration of TBG should be chosen that is high enough to bind a substantial amount of FITC-T4 before the addition of competitors (Huang 2003), but is low enough to avoid a large pool of free binding protein available for binding of competitors. Therefore, a concentration of 10 nM TBG was chosen.
T4 had a higher potency to displace FITC-T4 from TBG than from TTR (Fig. 4). The IC50 value of T4 with TTR was 125 nM and that of T4 with TBG 14.9 nM (Fig. 4, Table 3). Correspondingly, the estimated Ki value of T4 with TTR (41 (CI 34–49) nM, N = 3, n = 3) was determined to be approximately one order of magnitude higher than that of T4 with TBG (2.5 (CI 2–3) nM, N = 5, n = 2). A higher binding affinity of T4 to TBG than to TTR was also reported and discussed by Ren and Guo (2012) who attributed this to structural differences and different properties of the ligand binding pockets of TTR and TBG.
Fig. 4
Concentration–response curves of T4 in the TTR-binding assay (left y axis, N = 3, n = 3) and in TBG-binding assay (right y axis, N = 5, n = 2). Data are presented as the average (± SD) of separate experiments
Seven model compounds tested in the optimized TTR-binding assay and in the newly developed TBG-binding assayTo validate and compare the optimized TTR-binding assay and the newly developed TBG-binding assay methods, seven model compounds were selected and tested in both assays. Based on their previously reported competitive TTR binding the seven selected compounds were: T4 (as a positive reference chemical), D-mannitol (as a negative control), TBBPA, PFOS, PFOA, triclosan, and BPA (Collet et al. 2020; Weiss et al. 2015). The concentration–response curves of these compounds in the competitive TTR-binding assay are shown in Fig. 5a. Except for D-mannitol, all compounds displaced FITC-T4 from TTR with varying potencies (Table 3). TBBPA appeared to be the most potent compound (IC50 = 22 nM), and showed even a lower IC50 value than the endogenous thyroid hormone T4 (IC50 = 125 nM). The order of the competitive binding potency of the model compounds for TTR is: TBBPA > T4 > PFOS > PFOA > triclosan > BPA > > > D-mannitol. The corresponding IC50 and Ki values are shown in Table 3.
Fig. 5
Concentration–response curves for seven model compounds a tested with 110 nM FITC-T4 and 30 nM TTR at RT (N = 3, n = 3); b tested with 5 nM FITC-T4 and 10 nM TBG at RT (N = 5, n = 2 for T4 and N = 3, n = 3 for other model compounds). Data are presented as the average (± SD) of data from separate experiments
Figure 5b shows the concentration–response curves of the seven model compounds in the competitive TBG-binding assay. The negative control D-mannitol shows no inhibition of the FITC-T4 binding to TBG. No (or only limited) inhibition was observed for both PFOS and PFOA. Due to restricted solubility of both compounds in the assay medium, higher concentrations could not be tested. The model compounds showed displacement of FITC-T4 from TBG in the following order: T4 > triclosan > TBBPA > BPA > > > PFOA ≈ PFOS ≈ D-mannitol. The corresponding IC50 and Ki values are shown in Table 3.
Evaluation of the assays and comparison of outcomes with the seven model compoundsThe goal of the present study was to simplify and optimize the TTR-binding assay and develop a TBG-binding assay allowing easy, cheap and fast screening of chemicals for their binding capacity to TH distributor proteins. For both assays, similar Ki values of T4 were observed in the present and previous studies (Table 3). Results with seven model compounds showed that TBBPA, triclosan, PFOS, PFOA, and BPA compete with T4 for binding to TTR, which is in line with the results published by others (Cao et al. 2011; Cavanagh et al. 2018; Ren et al. 2016, 2020). In addition, the calculated Ki values of TBBPA, triclosan and PFOS are comparable to values published by others (Hamers et al. 2020), indicating that the updated experimental conditions (15 min, RT) do not affect the binding ability of the competitors to TTR to a great extent (Table 3).
According to different competitive binding experiments in the literature (Chi et al. 2020; Hamers et al. 2006) and this study, TBBPA binds more strongly to TTR than TTR’s natural ligand T4. While this is not the case for TBG, where T4 binds more strongly than TBBPA. Moreover, in the competitive TBG-binding assay, TBG demonstrated even a lower binding affinity (i.e., higher Ki) for TBBPA than TTR did (Table 3). Another FP-based competitive FITC-T4 binding assay with TTR and TBG showed binding of TBBPA to TTR, but not to TBG (Ren et al. 2020). In that study, the highest tested concentration was 100 µM TBBPA with 100 nM TBG and 50 nM FITC-T4 (Ren et al. 2020), whereas up to 250 µM TBBPA with 10 nM TBG and 5 nM FITC-T4 were used in the present study. These differences most probably result in a somewhat more sensitive screening, and therefore an effect of TBBPA in the TBG-binding assay in the present study.
BPA and its analogues have been reported to disturb the thyroid hormone system (Kim and Park 2019). BPA was the least potent chemical among the tested positive model compounds in both the TTR and the TBG assay (Fig. 5). Ki values of BPA for TTR- and TBG-binding derived from binding affinities reported by Cao et al. (2011) were one magnitude lower than those obtained in the present study (Table 3), indicating a weaker binding in the present study. In another study, however, Marchesini et al. (2006) observed no competitive binding to both TTR and TBG at the highest concentration of BPA tested (10 µM).
Triclosan was also reported to bind to TTR by using the ANSA probe and was also determined to be a more potent binder to TTR than BPA (Cavanagh et al. 2018). Few studies have investigated the binding capacity of triclosan with TBG. In a surface plasmon resonance biosensor assay with T4-coated chips and TBG, Marchesini et al. (2008) reported an IC50 value for triclosan of 1182 nM and for T4 of 17.2 nM. IC50 values from the present study were 1444 nM and 14.9 nM, respectively, being in line with the previous study. Direct comparison is difficult due to differences in test conditions and readout methods. Ki values for triclosan binding to TBG cannot be compared to previous studies, as to the best of our knowledge such Ki values have never been reported.
In agreement with previous findings by others, PFOS and PFOA bind to TTR but not to TBG (even when concentrations up to 1 mM were tested) (Ren et al. 2016). Most likely, PFOS and PFOA cannot adequately occupy the T4 binding pocket of TBG (Ren et al. 2016). Different binding affinities for TTR and TBG were not only reported for the compounds tested in the present study, but also for other compounds, including metabolites and derivatives of hydroxylated polychlorinated biphenyls (OH-PCBs) that bind stronger to TTR than to TBG (Cheek et al. 1999). These differences between TTR and TBG are most likely the result of differences in the binding pockets of TTR and TBG, i.e., hydrophobic channel T4 binding pockets in TTR and a surface binding pocket in TBG (Cao et al. 2011; Refetoff 2023). Hydroxylation, halogenation and presence of a hydroxylated phenyl ring has been reported to play crucial roles in the binding of chemicals to TH distributor proteins (especially the presence of an hydroxylated phenyl ring for binding to TBG) (Cao et al. 2010; Lans et al. 1994; Ren et al. 2016). This is confirmed by the data of the present study, as TBBPA, BPA, and triclosan all have these features (Fig. 1), while PFOS and PFOA miss a hydroxylated phenyl ring and do indeed not compete with T4 for binding to TBG. However, perfluorotridecanoic acid (PFTA) and perfluorotetradecanoic acid (PFTdA) that also lack a hydroxylated phenyl ring, were reported to bind to TBG with Ki values of 23 µM and 26.6 µM, respectively, which was explained with help of molecular docking that indicated that these longer fluorinated carbon chain acids fit the TBG binding pocket better than PFOA (Ren et al. 2016).
For the TTR-binding assay, the physiological relevance of the observed IC20 values for the model compounds was explored by a comparison with reported concentrations in human blood (Table S3). The TTR-binding capacity of a mixture consisting of the highest concentrations reported for the five TTR-binding model compounds in European biomonitoring studies was estimated to be equivalent to 19.1 nM of T4, according to the principle of concentration addition. The contribution of the different compounds to this T4-equivalent (T4EQ) concentration decreased in the order PFOS (50.9%) > triclosan (22.2%) ≈ TBBPA (19.5%) > PFOA (6.6%) > BPA (0.8%) (Table S3). Although the estimated 19.1 nM T4EQ only causes a 16% reduction of FITC-T4 binding to TTR in the bioassay, it is based on no more than concentrations of 5 xenobiotics in human blood. In a similar exercise with mixtures composed of maximum concentrations in human blood reported for 21 xenobiotics, the T4EQ concentration exceeded the IC20 in the assay by a factor of 3.5 (Hamers et al. 2020). In the same study, a mixture composed of median concentrations reported in human blood for the same 21 xenobiotics corresponded to 20% inhibition of FITC-T4 binding to TTR in the assay. Thus, the concentrations in human blood reported for individual xenobiotics may have a low TTR-binding capacity in the assay, but the combined mixture concentration of all TTR-binding xenobiotics in human blood gives a significant response. Similar estimation was also conducted based on the results of the TBG-binding assay (Table S4). The highest reported TBBPA, triclosan and BPA concentrations in blood in European countries were expressed in T4 equivalents, amounting to 2.2 nM of T4 equivalents, which is too low to displace T4 from TBG according to the concentration–response curve of T4. It must be noted, however, that one should be cautious when comparing nominal in vitro concentrations with total human blood concentrations, as there may be significant differences in concentration available for interaction with the targets (TTR and TBG) in vitro compared to in vivo. On the other hand, it should be realized that TTR is not only important as a distributor protein for TH in the blood, but also as a carrier protein for transporting TH across physiological barriers like the placenta or the blood-cerebrospinal-fluid barrier.
Impact of concentration–response analysis on Kd, Ki and IC valuesAs mentioned in “TBG-binding assay”, different approaches were taken to estimate Kd and Ki values in TBG-binding assays. The first approach was a descriptive model. Curve fitting was done using Eq. 1 to estimate Kd and Eq. 2 to estimate a maximum and minimum anisotropy, the IC50, and the Hill slope in Graphpad. The second approach was based on a theoretical model that took into account the principles of fluorescence polarization and of biochemistry with respect to protein binding. Kd values were obtained by the same principle as the first approach. Ki values, however, were estimated by solving the model numerically in R, and were then converted into ICX values. The idea behind taking two approaches was that the latter has a profound theoretical background, whereas the first is relatively easy to apply for users that are not experts in mathematical modelling, but rather prefer the use of vendor software like Graphpad. The equations used in theoretical model are described in the Supplementary Information file (Section 5) and the results obtained from the model are also shown in the Supplementary Information (Tables S2 and S5). The theoretically calculated Kd values were determined at 0.5 (CI 0.1–0.8) nM (22 °C) and 1.3 (CI 0.4–2.2) nM (37 °C) for temperature dependence experiments, and 1.1 (CI 1.0–1.2) nM at RT based on the repeated saturation experiments at RT (Table S2). As expected, these values are exactly the same as determined in Graphpad, because both methods use the same equation with least-squares fitting. Ki values estimated between both approaches were similar (Table S5).
留言 (0)