
記住我
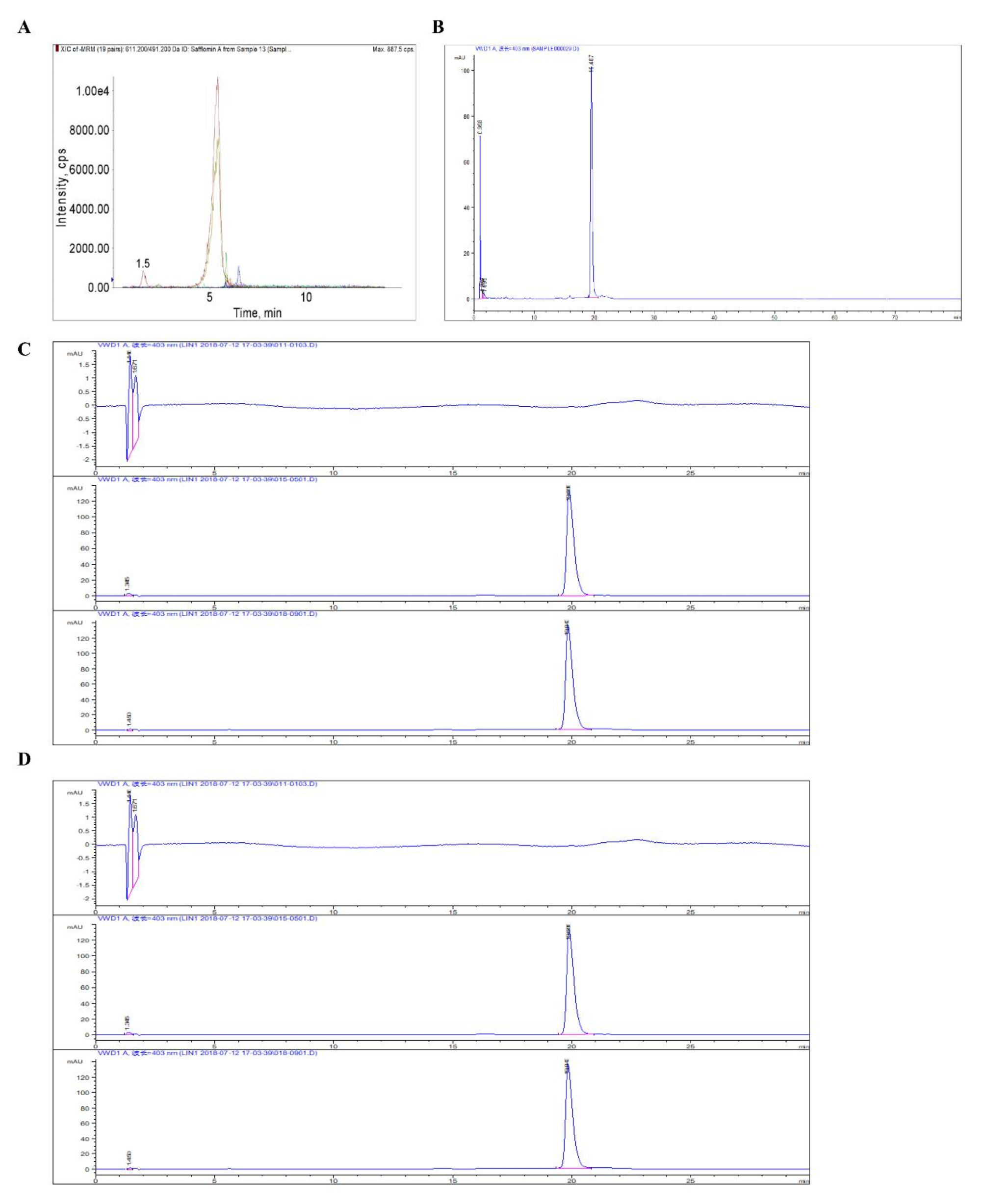





In total, 100 predicted DAU targets associated with drug absorption, distribution, metabolism, and excretion (ADME) were obtained from the SwissADME database, and 3036 AD-related targets from the GeneCards database. Intersecting these datasets yielded 66 common targets (Fig. 1A). PPI network analysis revealed a highly interconnected network with 66 nodes (proteins) and 291 edges (interactions), showcasing an average node degree of 8.82 and a local clustering coefficient of 0.6 (Fig. 1B). Further analysis of STRING-generated protein enrichment tables in Cytoscape 3.8.2 identified 16 core targets from PPI results (Table 2).
Fig. 1
Targets identification and network analysis of DAU for anti-AD activity through network pharmacology. (A) Venn diagram depicting common targets of DAU and AD; (B) PPI network of core targets. (C, D) GO function and KEGG pathway enrichment analysis. (E) Predicted compound-target-biological process-pathway network
Table 2 Core targets screened by Cytoscape based on PPIFunctional enrichment analysis identified 21 significantly enriched GO terms, including 13 biological processes (BP), 4 cellular components (CC), and 4 molecular functions (MF) terms (Fig. 1C). Prominent enriched BP terms included platelet activation, positive regulation of MAP kinase activity, peptidyl-threonine phosphorylation, protein kinase activity activation, peptidyl-serine phosphorylation, and protein autophosphorylation. Enrichment in CC implicated the plasma membrane, internal component of the plasma membrane, dendrite, and postsynapse. MF terms encompassed kinase activity, enzyme binding, protein kinase activity, and protein serine/threonine kinase activity. These findings suggest DAU primarily targets the cytoplasmic membrane, particularly with relevance to neurosynaptic function, and likely exerts its effects through kinase-level regulation. KEGG pathway enrichment analysis identified 17 potential pathways, involving neuroactive ligand-receptor interaction, estrogen signaling, thyroid hormone signaling, Rap1 signaling and so on (Fig. 1D). According to P-value and gene number analysis, the most relevant is the neuroactive ligand-receptor interaction pathway.
Network analysis is used to predict the central target with the best degree of connectivity. Compound-target-biological process-pathway network analysis showed that DAU interacted with multiple potential targets and signaling pathways, highlighting potential synergistic therapeutic effects (Fig. 1E). Based on the degree values in the node table in the network graph, we selected the top ranked targets (Table 3). Considering the relevance of the target and the disease, as well as the pertinent research background, we speculate the most potential mechanism of DAU’s anti-AD effect is related to the regulation of PIK3CA, AKT1 and mTOR.
Table 3 Connectivity of core targets from network of “compound-target-biological process-pathway”Molecular docking simulation analysisTo validate network pharmacology predictions and elucidate the potential binding interactions, molecular docking simulations were performed between DAU (ligand) and the three core targets, PIK3CA, AKT1 and mTOR (receptors). Lower binding energy indicates more stable interaction [26]. DAU exhibited strong binding affinity towards PIK3CA, AKT1 and mTOR, with binding energies of -10.4, -6.8, and − 7.7 kcal/mol, respectively. These findings suggest that these proteins may hold significant therapeutic potential for DAU against AD. Additionally, Fig. 2 visually demostrated the formation of stable DAU-target complexes, further highlighting the reliability of network pharmacology predictions.
Fig. 2
Three-dimensional (Left) and two-dimensional (Right) docking models of DAU binding to PIK3CA, AKT1 and mTOR
Alleviation of Aβ-induced paralysis by DAU in AD nematodesTo investigate the in vivo anti-AD activity of DAU, AD transgenic nematodes CL4176 and CL2006 were used. These nematodes express human Aβ protein in their body wall muscle cells and exhibit paralysis phenotype at different times as the protein aggregated (CL2006 at 20 °C for ~ 10 days, CL4176 rapidly entered paralysis upon temperature induction). Before the examination, food clearance assay was firstly taken to determine the safe dosing range of DAU in wild type and AD modle nematodes. Results shown in Fig. S1, revealed no influence of DAU treatmen on the growth and development of nematodes by the dose concentration below 80 mM. Considering the concentration of DAU in previous trial studies, we opted for the administration of DAU ≤ 10 µM.
Paralysis assay of CL4176 nematodes were shown in Fig. 3A. DAU treatment significantly shifted the paralysis inhibition curves upwards, and demonstrated an anti-paralytic effect in CL4176 (P value shown in Table S1). This observation was supported by the PT50 values (the time to reach half-paralysis) in Fig. 3B, with a paralysis time delay from 4 h (Control) to 6 h (5 µM, P = 0.006) and 8 h (10 µM, P = 0.0001). The 10 µM DAU group exhibited the longest PT50, demonstrating superior paralysis inhibition efficacy. This optimal concentration was used for subsequent experiments. Furthermore, administration of 10 µM DAU also exhibited a significant anti-paralysis effect in CL2006 (Fig. 3C), reducing the PT50 from 6 h to 10 h (Table S2, P < 0.0001), indicating an alleviation of Aβ-induced toxicity. These findings collectively suggest the promising in vivo anti-AD activity of DAU.
Fig. 3
Effect of DAU on Aβ-induced paralysis in CL4176 and CL2006. (A) Non-paralysis curves diagram of CL4176; (B) The mean of time at which 50% CL4176 worms were paralyzed (PT50) with different doses of DAU. (C). Non-paralysis curves diagram of CL2006 with 10 µM DAU. Results are representative of three independent experiments and presented as mean ± SD. *P<0.05
Target-based activation of autophagy by DAU in C. elegansIn CL4176 nematodes, DAU significantly downregulated the expression levels of C. elegans homologs, age-1 (PIK3CA), akt-1 (AKT1), and let-363 (mTOR) (Fig. 4A), validating network pharmacology predictions. Since PI3K/AKT/mTOR signaling pathway regulates autophagy, a key process critical for clearing Aβ aggregates, we investigated DAU’s influence on the expression of autophagy-related genes (lgg-1, bec-1, unc-51, atg-18, and epg-8) in CL4176 and CL2006 strains. Results shown in Fig. 4B, C revealed that DAU treatment significantly upregulated these genes, indicating potential autophagy activation. This was further supported by the increased GFP-positive punctae (13 to 26) in DA2123 nematodes expressing the gfp::lgg-1 autophagy reporter gene after DAU administration (Fig. 4D).
To further validate the correlation between autophagy activation and DAU’s anti-AD effect, we co-administered DAU with the autophagy inhibitor 10 µM 3-methyladenine (3-MA, MCE, New Jersey, USA), which primarily inhibits Class PI3K-III activity and prevents autophagosome formation and LC3 expression [27]. Results were shown in Fig. 4E. DAU’s anti-paralysis effect (red curve, P = 0.0003) was nullified upon 3-MA co-treatment (blue curve, P = 0.3643), while the PT50 were induced from 7 h to 4 h (Table S3), indicating that DAU’s beneficial effect on the inhibition of Aβ-induced paralysis is associated with autophagy activation, particularly autophagosome formation.
Fig. 4
Effect of DAU on autophagy activation. (A) Core target genes expression in CL4176 nematodes; (B) Autophagy genes expression in CL4176; (C) Autophagy genes expression in CL2006; (D) Representative micrographs of GFP-positive punctae in DA2123 carrying the gfp::lgg-1 reporter gene (Left, Scale bar: 50 μm) and quantification of GFP-positive punctae per worm with/without DAU (Right); (E) Non-paralysis curves diagram of CL4176 with 3-MA co-treatment. Results are representative of three independent experiments and presented as mean ± SD. * P < 0.05; ** P < 0.01
Facilitation of autophagic lysosomal fusion and degradation by DAU in C. elegansEfficient fusion of autophagosomes with lysosomes and subsequent degradation are crucial for autophagy completion and efficacy. To investigate DAU’s impact on this process, we assessed lysosomal content and autophagic substrate consumption. Lyso-Tracker red dye staining revealed significant increased lysosomal activity in DAU-treated worms compared to the control group (Fig. 5A). Then, we utilized BC12921 strain, expressing GFP-tagged autophagic substrate protein P62, to monitor autophagic substrate consumption. DAU treatment significantly reduced GFP fluorescence intensity (Fig. 5B), indicating increased autophagy degradation activity and autophagic substrate consumption.
To investigate the correlation between DAU’s autophagic degradation activity and its anti-AD efficacy, we co-administered DAU with 100 µM chloroquine (CQ; MCE, New Jersey, USA), an inhibitor of autophagosome-lysosome fusion. DAU’s ameliorative effect (red curve, P = 0.0005) against CL4176 paralysis disappeared after co-treatment with CQ (purple curve, P = 0.4125) (Fig. 5C, Table S4), indicating that DAU’s anti-AD efficacy was associated with the activation of autophagic lysosomal fusion and degradation.
Fig. 5
Effect of DAU on autophagic lysosome fusion and degradation in nematodes. (A) Lysosome content revealed by Lyso-Tracker red staining in CL4176. Scale bar: 50 μm; (B) Expression level of autophagic substrate protein P62::GFP in BC12921. Scale bar: 50 μm; (C) Non-paralysis curves diagram of CL4176 co-treated with DAU and CQ. Data represent mean ± SD from three independent experiments. * P < 0.05, ** P < 0.01
Exclusion of autophagy activation by DR in C. elegansDietary restriction (DR) has long been associated with the induction of autophagy and the reduction of proteotoxicity in C. elegans and other animals [28]. In order to confirm whether the activation effect of DAU on autophagy is related to DR, we examined the influence of DAU on the fat accumulation and the feeding behavior in C. elegans N2. The ORO staining was used to evaluate the effect of DAU on the fat accumulation in nematodes. As shown in Fig. 6A, B, the quantification of staining intensity in glucose-treated worms (high-fat positive control) was significantly higher than control, while the DAU-treated nematodes exhibited no difference with control, indicating no influence of DAU on the lipid content in C. elegans and no nutritional deficits exist. Nematode pharyngeal pumping rate was examined to reveal the food intake influenced by DAU, and the results were shown in Fig. 6C. Compared to control, neither day 3 nor day 6 after DAU treatment, there was a significant difference on the pumping number, indicating no effect of DAU on food intake. Therefore, we speculated that the activation effect of DAU on autophagy in nematodes was independed on DR.
Fig. 6
Effect of DAU on the fat accumulation and pharyngeal pump rate in wild type N2 nematodes. (A) Representative micrographs of ORO staining of worms treated with Glucose (positive control), DMSO and DUA (Scale bar: 100 μm). (B) The quantification of ORO staining intensity per nematode of different groups. (C) Pumping number per nematode in 30 s measured on the third and sixth day after DAU treatment. Data are presented as mean ± SD from three independent experiments. ** P < 0.01
Alleviation of Aβ aggregation by DAU in C. elegansThe pathogenicity of Aβ aggregation stems from the abnormal accumulation of Aβ proteins, and mitigating this aggregation can significantly alleviate the associated toxicity [29]. We investigated the potential correlation between DAU’s anti-AD efficacy and Aβ clearance by assessing Aβ accumulation in CL4176 and CL2006 strains using Th-T staining. Th-T dye exhibits high affinity for Aβ aggregates, enabling quantification of protein aggregation through fluorescence intensity in the anterior pharynx, a key Aβ deposition site in these models. DAU treatment markedly reduced Aβ deposits in the anterior pharyngeal region of both CL4176 and CL2006 strains compared to controls (Fig. 7A, B), suggesting it effectively eliminates abnormal aggregation of Aβ proteins, primarily through autophagy-lysosome pathway.
Fig. 7
Effect of DAU on Aβ aggregation in CL4176 (A) and CL2006 (B) strains by Th-T staining. (Left) Representative micrographs of Aβ aggregation treated with or without DAU. White arrows indicated Aβ aggregation sites with the scale bar of 50 μm. (Right) Statistics on the number of Aβ deposits per nematode in different groups. Error bars represent the mean ± SD. * P < 0.05
Inhibition of Aβ toxicity by DAU at different administration time of C. elegansThe impact of drug administration timing on diseases is of significant importance in elucidating the mechanism of drug action and its medicinal application. The aforementioned results indicate that prior treatment of DAU before Aβ induction at L1 larvae holds promising potential as an effective prophylactic medication for AD. In order to explore whether DAU has the same anti-AD effect after the onset of disease, we investigated the neuroprotective capacity of DAU at 12 h/ 24 h after Aβ induction (23 °C) using C. elegans CL2355, which contains pan-neuronal Aβ expresion and deficits in chemotaxis behavior [30]. As shown in Fig. 8A, B, compared to the blank strain CL2122 (no Aβ expression), the chemotaxis indexe of CL2355 was decreased from 0.48 to 0.25, demonstrating the neurological impairment in perceiving the attractant benzaldehyde. After DAU administration, no matter when the drug treated before (L1 larvae) or after Aβ induction (12 h/ 24 h), the chemotaxis indexes of DAU groups were significantly increased to 0.43, 0.36 and 0.34 compared to the untreated CL2355. This result confirmed the protective effect of DAU at older ages of AD nematodes, indicating a persistent benefits of autophagy activation to potential therapeutic efficacy of DAU.
Fig. 8
Effect of DAU on Aβ-mediated behavioral dysfunction in CL2355 nematodes. (A) Schematic diagram of the chemotaxis assay plate. (B) Chemotaxis indexes of CL2355 with DAU administration at different time (L1- before Aβ induction; 12 h/24 h- after Aβ induction). Approximately 300 nematodes were analysed for each treatment and the results are presented as means ± SD of three replicates. *** P < 0.001, **** P < 0.0001
Clearance of polyQ proteins by DAU in C. elegansPathogenic protein aggregation is a major molecular mechanism in protein conformational diseases, including AD and HD. We explored DAU’s potential to scavenge polyglutamine (polyQ), the pathogenic protein of HD, using a HD model strain AM141. This strain expresses polyQ40::YFP fusion proteins in body-wall muscle cells, resulting in discrete fluorescence aggregates over time (Fig. 9A) and malfunction of motility. Results were shown in Fig. 9. Although early DAU treatment (48 h, 72 h) didn’t significantly impact polyQ aggregation and toxicity, it markedly reduced the number of fluorescent aggregates in AM141 body wall (Fig. 9B) and improved the rate of body bends at 96 h post-DAU administration (Fig. 9C). These findings suggest that DAU can reduce toxic protein PolyQ aggregation, which should be also related to the activation of autophagy-lysosome protein degradation pathway by DAU.
Fig. 9
Effect of DAU on PolyQ aggregation and motility in AM141 nematodes. (A) Fluorescence microscopy images of AM141 nematodes at 48, 72, and 96 h post DAU treatment. Scale bars: 100 μm. (B) Quantification of polyQ40::YFP aggregates. (C) Body bends number of AM141 per min. Data present mean ± SD from three independent experiments. ** P < 0.01
留言 (0)