
記住我
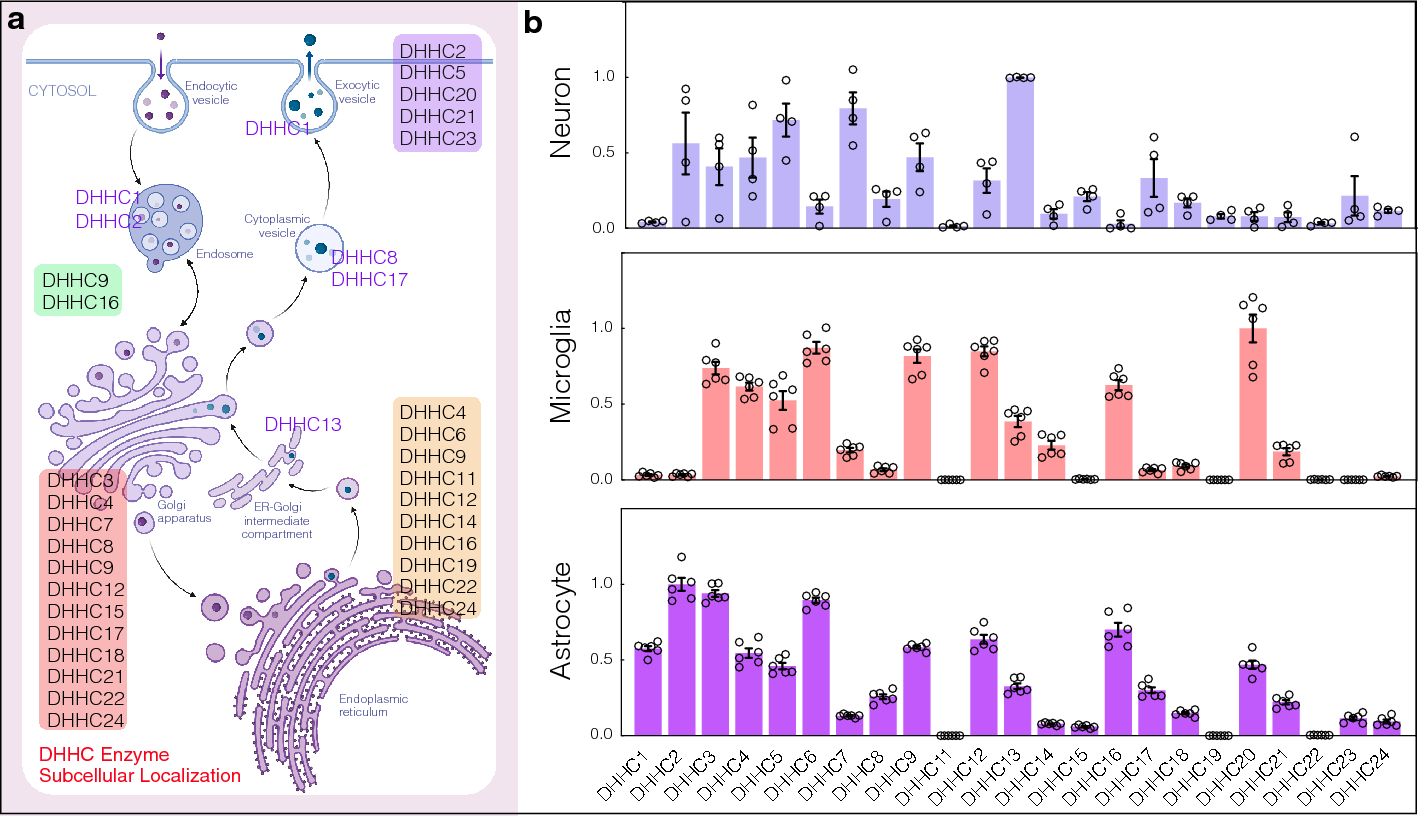




BEEF-CP is an analog of the common Ca2+-sensitive dyes Calcium Green and Oregon Green that features nonpolar and electron-donating substituents, namely ethyl groups instead of halogen substituents on the fluorescein moiety (Fig. 1a). Fluorescein-based fluorophores are well known for their nonfluorescent lactone and fluorescent zwitterion tautomerism, which is heavily influenced by the substituents on the xanthene ring, pH, and other factors [26]. At the same time, the pH and Ca2+-dependent fluorescence of BAPTA-based Ca2+ indicators are a key factor in their applicability. At low or high pH values, these dyes can give false negative or false positive signals, respectively. The Ca2+ turn-on fluorescence experiments of BEEF-CP demonstrated low background and high enhancement in the biologically relevant pH range of 6.0–8.0 and consequently confirmed its suitability for in vitro measurements (Supplementary Fig. 21). The use of BEEF-CP in uninfected VeroE6 cells in a concentration of 0.5 µM was not associated with any cell toxicity signs measured over 24 h in the biosafety laboratory where these measurements were run along with cell infection experiments.
Fig. 1
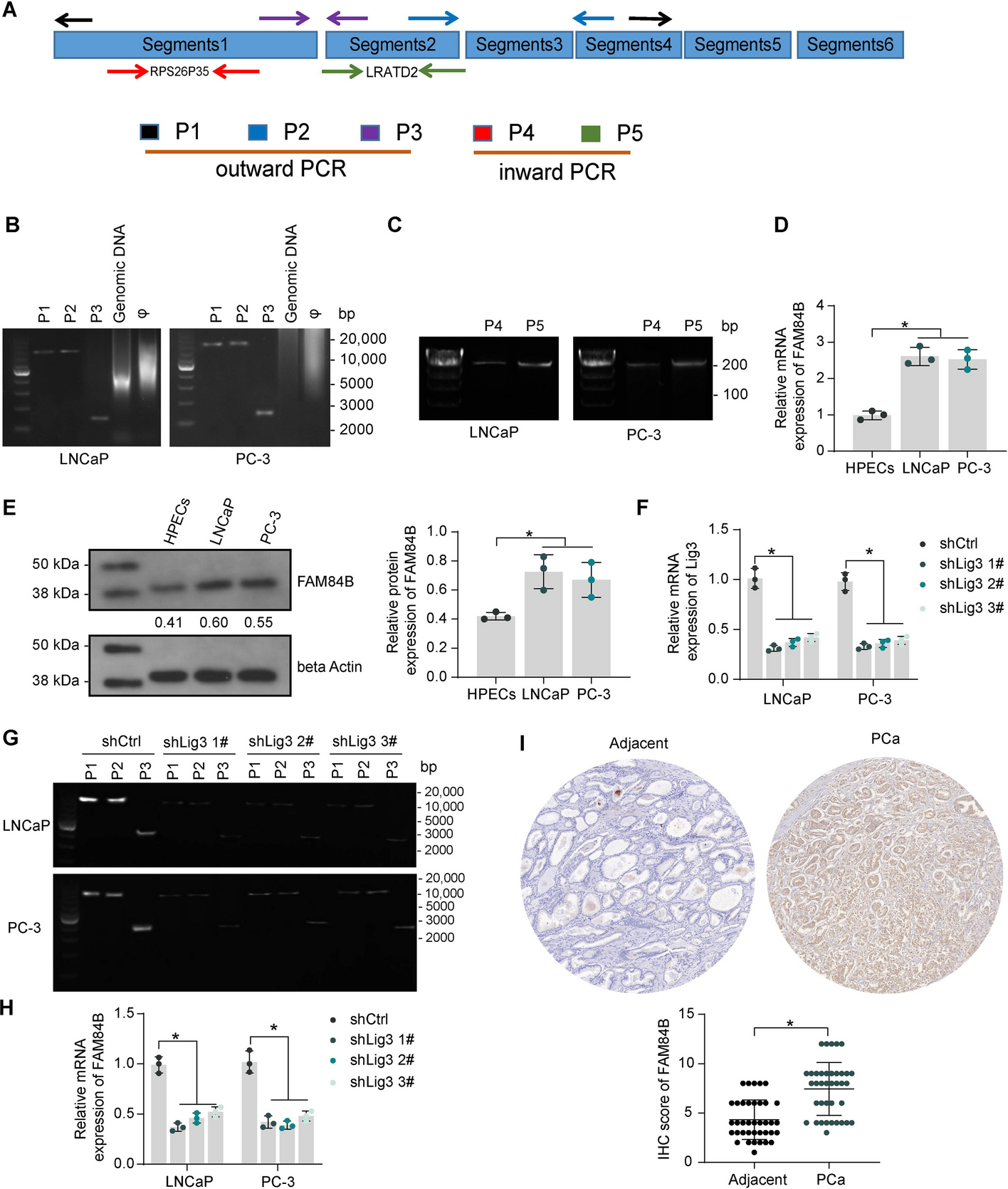
Experimental design and manual analysis of infection in Vero E6 cells using SARS-CoV-2 variants D614G and B.1.1.7. A Fluorescence enhancement of the BEEF-CP dye upon Ca2+ binding is attributed to the alleviation of the photoinduced electron transfer (PET) quenching. Its low background fluorescence and high two-photon cross-section (TPCS) along with its advantageous cell internalization make BEEF-CP an appropriate candidate for cytosolic Ca2+ imaging with 2P microscopy. B Experimental design with 2P microscopy. A pulsed Ti:Sa laser source was used for excitation at 700 nm wavelength, and laser light was diffracted with a pair of galvo mirrors for scanning the region of interest. A high-numerical-aperture objective (Olympus 20×) was used to obtain subcellular resolution. Samples were prepared in 96-well bioassay plates and transferred directly to the microscope. Samples were moved semi-automatically to locate the different wells in the field of view of the microscope. C Exemplary images from samples where most cells correspond to a particular stage of the infection (see “Methods” for more details on the characterization criteria). D The relative number of cells in different stages of infection 48 h post infection for the two variants at two different dye concentrations at various virus titers. Data from wells with the same virus titer, variant, and dye concentration are averaged. Statistical data for the individual cell categories are shown in Supplementary Fig. 5. E 50th percentile values, i.e., the percentage of cells under the median severity of infection, for the data displayed in D. Statistical analysis of the manual cell sorting shows that (i) the ratio of healthy cells is significantly higher with lower virus titer, regardless of the virus variant or dye concentration; and (ii) for high infection levels (TCID50 > 10–3 mL−1), the proportion of absent cells is higher for the D614G variant
The spectroscopic characterization of BEEF-CP involved one-photon (1P) and 2P fluorescent measurements and the quantum yield (QY) determination in the presence and absence of Ca2+ ions. The measured fluorescence intensity at pH 7.2 showed a single-step enhancement upon Ca2+ addition in accordance with the formation of a single monometallic complex. The fluorescence enhancement was found to be around threefold in the biologically relevant Ca2+ concentration range, from 10 nM to 1 µM (Supplementary Fig. 20). The dissociation constant of the Ca2+ complex (KD = 175.6 nM) is comparable with previously synthesized molecular probes for in vivo calcium level determination [34, 35]. The background fluorescence of BEEF-CP is not zero in the absence of Ca2+, which provides appropriate contrast to visualize even the healthy cells with a low level of Ca2+ ion during the cell viability assay. Furthermore, the high-slope region of the Ca2+-dependent fluorescent curve is adequately positioned to distinguish between healthy and malfunctioning cells.
The further applicability of BEEF-CP in cellular systems was tested by cytometry measurements on the HEK-293 cell line. Stained cells showed an increased mean fluorescence intensity by around one order of magnitude compared with the control level (Supplementary Fig. 17). In a parallel experiment, the stained HEK cells were treated with ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA) prior to the analysis, which resulted in a somewhat lower mean fluorescence intensity. In contrast, cells stained in the presence of ionomycin using an otherwise identical procedure exhibit substantially higher mean fluorescence intensity. The ionomycin releases exclusively the intracellular Ca2+ storage, which proves that the dye is internalized by the cells as it responds to the cytosolic Ca2+ levels [36] (Table S5).
Subcellular dye localization is characteristic and reproducibly different in uninfected and infected cell culturesTo perform high-resolution quantitative analysis of the level of SARS-CoV-2 infection, we used 2P imaging. This imaging technique has a high, subcellular spatial resolution (~ 350 nm) and virtually no out-of-focus signal from nonstudied cells or compartments, above or below of the imaging plane, which ensures that the detected signal originates only from the cells of interest. Samples were prepared on a 96-well bioassay plate with a clear, flat bottom for good optical access. One to three high-resolution, 2P, full-field, 360 × 360 µm large images were acquired with a Femto Smart 2P microscope at 700 nm wavelength (see Supplementary Fig. 3 for wavelength selection) from one or more representative area within each sample (see “Methods” for details).
Acquired images were analyzed with two complementary methods (manual and automatic). During manual evaluation, the cells were sorted according to their morphology (Fig. 1b–d), while the automatic analysis, based on statistical methods, consisted of the classification of the full images based on a set of image parameters (Fig. 2). Both methods take the different features of the acquired raw images into account for the analysis, and for both we investigated the fine details of the images (at cellular or single-pixel level) for designing a quantitative measure correlating with the initial challenge infection level. Two variants of concern of SARS-CoV-2, namely D614G and B.1.1.7, were studied (Supplementary Fig. 6). These caused pathophysiologically and clinically distinguishable disease patterns in the clinic, so the putative infection quantitation and infection assay properties of BEEF-CP would be tested as the distinguishing features of cellular Ca2+-related effects of these two variants. Different cellular features such as the state of infection and the apoptotic-like state were visible and distinguishable on the images of Vero E6 cells infected with the two SARS-CoV-2 variants (Fig. 1b). The investigations were done in a phase when still living cells (both healthy and infected) adhered to the plate surface. Cell compartments, such as cell membrane and nucleus, were visible and distinguishable in the samples (Supplementary Fig. 15). With increasing virus titer, the mean fluorescence of infected cells increased significantly as this caused more Ca2+ to enter the cytoplasm. When the infecting viral titer increased even further, the morphology of cells changed, and dead cells detached from the wells plate; thus, the number of these absent cells could be estimated by the ratio of unpopulated areas at the plates’ surface. Due to the subcellular resolution of this cell viability assay, we could detect the formation of several syncytial formations, as giant multinucleated cells in the most infected cultures, which agrees with the earlier observation that SARS-CoV-2 virus causes this type of cellular fusion (Supplementary Fig. 2) [37, 38].
Fig. 2
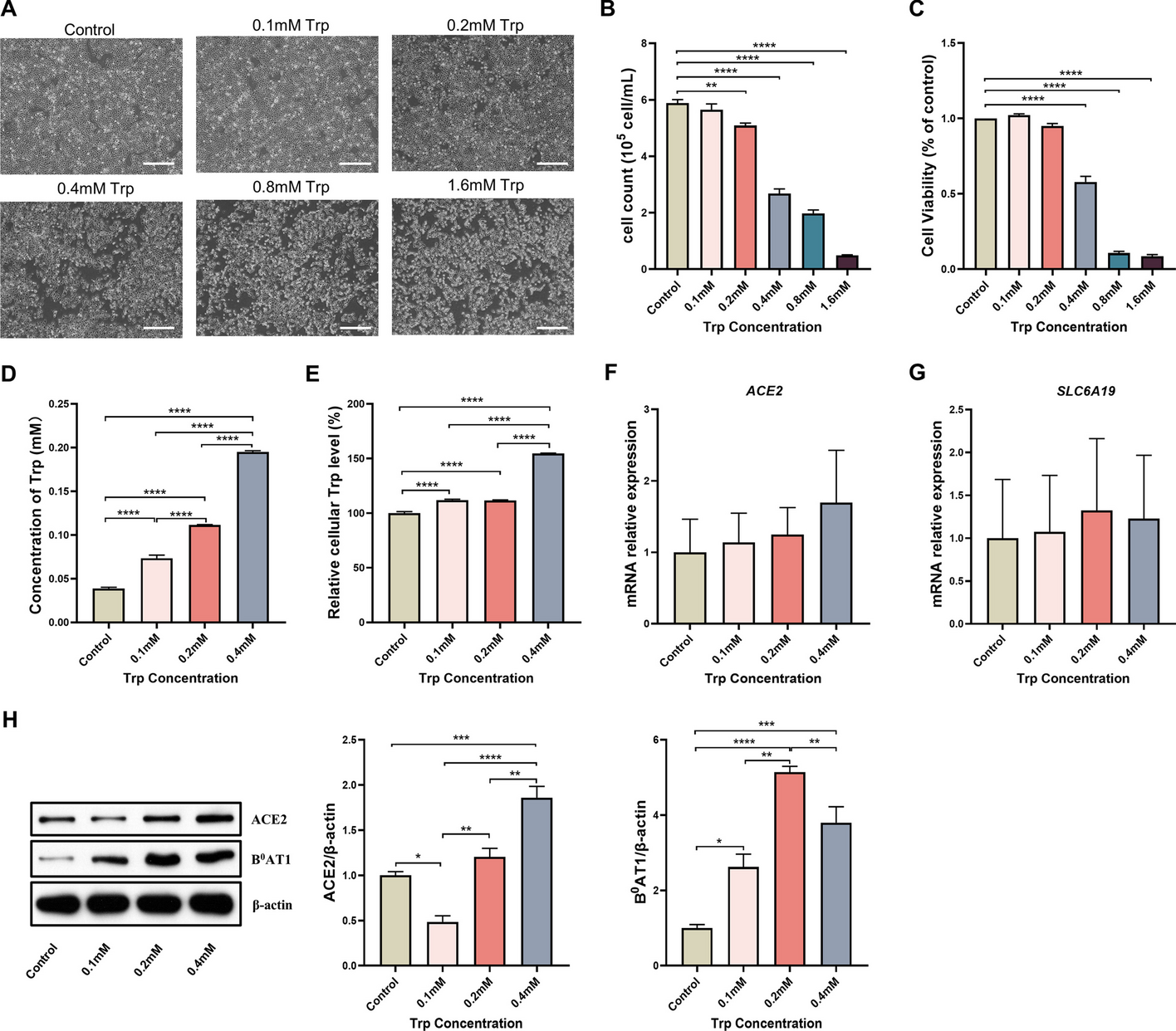
Automatic analysis of the 2P microscopy images of infected cells using SARS-CoV-2 variants D614G and B.1.1.7. A Purple contours indicate the areas automatically defined as particles during this analysis. B, C, E, F Spider charts showing the effects of B variant, C virus titer, E dye concentration, and F photomultiplier tube (PMT) relative voltage on the range-normalized values of seven different image parameters obtained from 2P microscopy images. The error bars for the spider charts can be found in Supplementary Fig. 7. D The image parameter called “relative signal area” shows a significant increase at virus titers higher than TCID50 10–3 mL−1. G The image parameter called “image mean intensity” for the two studied variants at high virus titer (TCID50 > 10–3 mL−1) at 0.05 μM and 0.5 μM dye concentration. H t-Distributed stochastic neighbor embedding (t-SNE) 2D plot obtained from all the seven image parameters recorded at 0.5 µM dye concentration shows three clusters depending on the virus variant and titer. Inset shows that the three groups, namely no or low infection, infection with D614G variant, and infection with B.1.1.7 variant are clearly separated along the first dimension. I Classification of the images corresponding to different variants at various virus titer in three clusters by seven-dimensional Gaussian mixture model clustering (error bars show standard deviation; significance levels as *p ≤ 0.1; **p ≤ 0.05; ***p ≤ 0.01.)
Manual cell counting quantifies infection progressDuring the manual evaluation, cells were manually localized on the recorded images, and then the individual cells were visually categorized into five groups, representing different stages of the infection (see Fig. 1c and “Methods” for more details on cell categories). The individual images from different wells were subjected to a single-blind evaluation by a human operator. The cells were classified into four categories (Fig. 1c), while the fifth category, cell death, was determined by the absence of living cells, estimated from the surface area that was not covered with cells, as shown in Fig. 1c. The number of dead cells is not directly counted; instead, it was estimated from the number of missing or absence of cells relative to an average number of cells from images without infections. However, one may observe some variability in the number of overall cells per field of view, provided in Supplementary Tables 1 and 2. The ratio of the cells belonging to each category was calculated for the different virus variants, virus titer, and dye concentration showing an increasing number of cells in the healthy or initial infection categories with the virus titer decreasing (Fig. 1d and Supplementary Tables 1 and 2). The 50th percentile values were calculated for each virus titer by averaging the result from all wells infected with the given titer. For this calculation, cells were sorted by the severity of their infection from healthy to absent dead cells, and the position of the median was calculated as the ratio of all the cells. The 50th percentile values also show a clear trend with increasing viral load. Furthermore, there is a clear jump in the severity of the infection in the cell culture between TCID50 virus titer 10–4 and 10–3 mL−1.
Automatic image analysis identifies advanced infection and virus variantAn ImageJ [39] macro was created to identify particle objects based on an intensity threshold (Fig. 2a) and to extract image data from the fluorescence microscopy images in the form of a few numerical parameters. The ImageJ macro is available in Appendix A. Seven parameters characterizing the detected particles were defined in each image: (i) relative signal area, (ii) image mean intensity, (iii) mean of the threshold area, (iv) maximum particle intensity, (v) average particle size, (vi) particle percentage area, and (vii) particle mean intensity (Supplementary Table 3).
The input images correspond to experimental points that can be described by two infection-related variables, namely virus titer and virus variant, and by two measurement-related variables, namely BEEF-CP sensor concentration and photomultiplier tube (PMT) voltage. Analysis of the image parameters as a function of the infection-related and measurement-related variables can yield information about the efficiency and robustness of the automatic image analysis, respectively. Figure 2b–c, e–f shows that the image parameters vary according to different acquisition settings, i.e., virus concentration, virus type, dye concentration, and PMT voltage. Importantly, certain image parameters such as “relative signal area” (Fig. 2d) can be a good indicator of the viral infection as they show increasing parameter values with increasing virus titer. However, images of the two variants did not show clear separation by any parameter values without eliminating the variation caused by the measurement-related variables.
Other prevalent Ca2+ sensors, such as OGB-1 AM (Oregon Green 488 BAPTA-AM), are used at a high concentration, around 1 mM (ranging from 0.9 to 1.4 mM) [40,41,42]. We used BEEF-CP at lower concentrations, at 0.5 µM and 0.05 µM. Both concentrations were suitable to differentiate between little or no infection and a high level of infection using automatic image analysis. Both these concentrations, however, are above the typical toxicity level, influencing the results. Moreover, differentiation of the variants responsible for the infection became possible when the images obtained with higher dye concentration (0.5 µM) were analyzed in those cases when the well was highly infected (TCID50 > 10–3 mL1) (Fig. 2g). Therefore, for further analysis, only those images were considered that were acquired from samples stained with 0.5 µM dye concentration.
A cluster analysis that considers all seven parameters was run on all the images acquired using 0.5 µM dye concentration. Two-dimensional visualization of the seven-dimensional clustering (Fig. 2h) shows three separated clusters (see Experimental procedures and Supplementary Figs. 7–14 for the detailed analysis). A three-component Gaussian mixture model cluster analysis was performed on the image parameters. Figure 2i shows that the three obtained clusters agree well with the three distinct states of the wells, i.e., little or no infection, highly infected with the D614G virus variant, and highly infected with the B.1.1.7 variant.
Quantitative image analysis shows a viral variant-dependent separation and a clear infection threshold of the applied virus titersThe manual and automatic analyses both showed a difference between the two tested virus variants, with D614G having a significantly higher infection rate. This finding complements earlier results that found a higher infection rate for D614G than B.1.1.7 in HeLa ACE2 and similar rates in HEK-293 T cells [43]. There is a very clear threshold found over 10–3 mL−1 TCID50 virus titer, where instead of occasional virus infection spread, we saw reliably escalated infection at all imaging sites. Furthermore, the automatic analysis revealed other important findings. First, it showed a clear threshold for the dye titer, showing that at least 0.5 µM dye concentration is required for the significant differentiation between variants (Fig. 2g). The results suggest that the fluorescence intensity of infected cells—related to the image mean intensity—is delimited by the dye concentration at 0.05 µM and by the Ca2+ concentration at 0.5 µM dye load. Second, the cluster analysis of the data could distinguish three separate groups (Supplementary Fig. 12). Among these was the noninfected group; the images falling into this cluster showed similar characteristics to the control measurements. However, more interestingly, the remaining images were almost perfectly separated by the virus variant used for the infection (Fig. 2h). These results demonstrate that this method is capable of quantifying the infection level and can also differentiate the two applied variants, D614G and B.1.1.7.
留言 (0)