For archiving the numerous mouse lines that are being constantly developed for biomedical research, freezing of sperm has become increasingly important compared to the archiving of pre-implantation embryos. In routine practice, sperm are obtained from the cauda epididymis of previously killed donor males. We have successfully tested a novel, non-terminal sperm collection method for mice by the separation of mating partners shortly before the onset of ejaculation. As sperm can be collected repeatedly with this approach, the method is also of interest for other scientific purposes. These include the assisted reproduction of a line by IVF or the assessment of the donor’s sperm quality over longer periods of time or at different stages of life. Currently, there is no other method to repeatedly collect sperm samples from mice without stressing the donors.
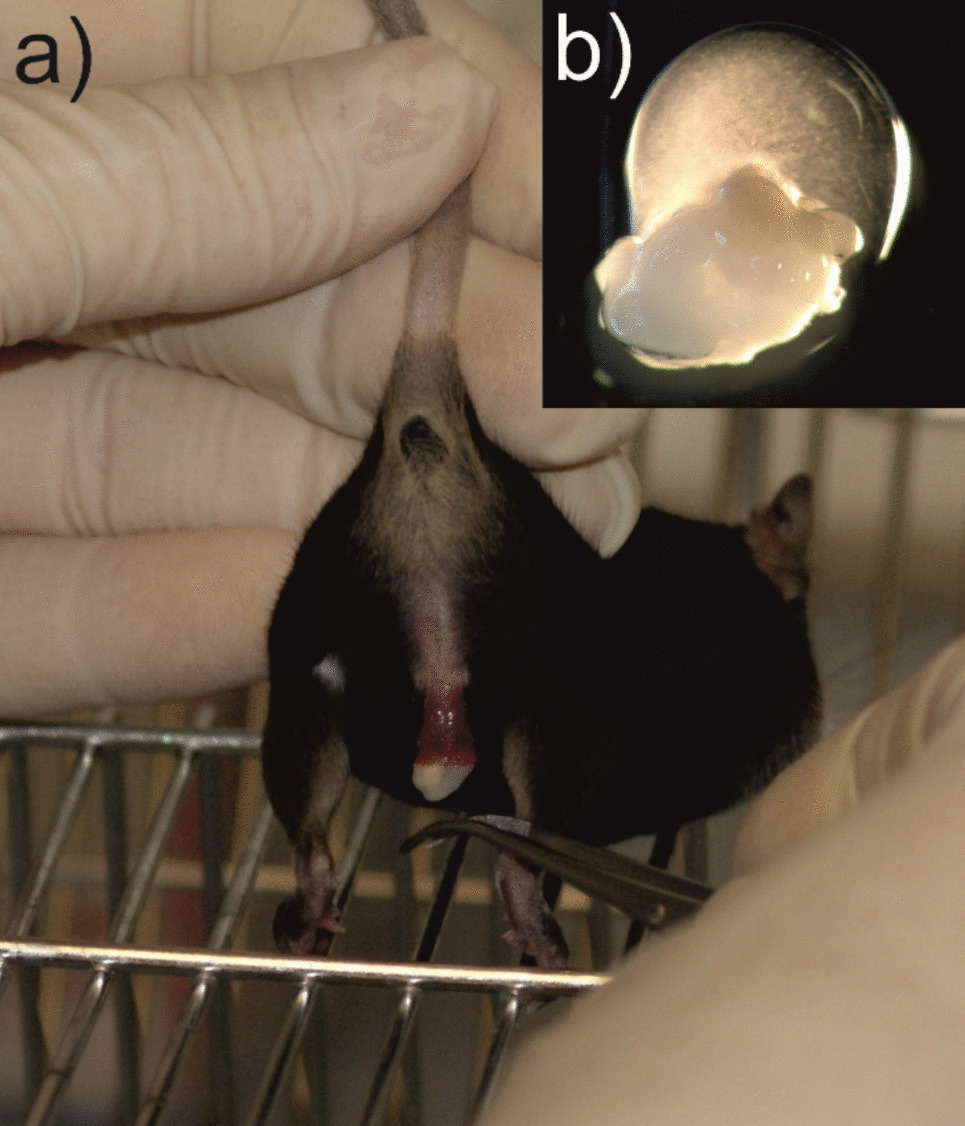
To obtain the ejaculate, the male has to be separated from the female within 1–2 s after the onset of the ejaculatory shudder. Recognizing the correct time point to separate and carrying out the separation can be learned quickly. It is worth noting that the success rate for the first ten ejaculations was only 10% but increased rapidly thereafter. After the first 20 ejaculations, the success rate in obtaining the ejaculate was on average over 90%. None of the females that were separated at the proper time and therefore had no vaginal plug became pregnant. If, however, separation would occur too late, the Bruce effect could be used to terminate the unwanted pregnancy by exposing the female to the pheromones of an unfamiliar male [26].
During the 11 to 14 matings per male, we did not observe a decrease in the males’ readiness to mate. This observation suggests that the separation just before ejaculation and the subsequent removal of the copulatory plug with the ejaculate from the penile cup does not inflict a stressful or otherwise adverse event for donor males.
The most time-consuming aspect of this sperm collection method was the search for a receptive female, as female behavior did not always clearly indicate acceptance or rejection of the male. On some scheduled test days, none of the used females could be identified as clearly receptive. Increasing the number of available females would have helped to overcome this problem, as we temporarily lost females from the pool due to pregnancies, especially at the beginning of the study. However, once found, an estrous female can be presented to several donor males in succession on the same day, given that the males are separated at the correct time point before ejaculation. Here, it was possible to obtain sperm from all ten males tested after mating with the same receptive female. Estrous females used also showed no decline in their readiness to mate with several males in succession, a behavior supported by observations of multiple mating and multiple paternity in Mus domesticus [27] and Mus musculus [28]. As an alternative to mating female mice, estrus cycle examination using vaginal swabs could be used to identify receptive females [29].
In an experiment with male offspring of pregnant wild-caught Mus m. domesticus females, the sperm count in mouse ejaculates was determined by flushing the reproductive tract of the female B6CF1 mating partners. The results showed a large variation between males, ranging from 3.48 × 106 to 16.53 × 106 sperm per recovered ejaculate [30]. The mean sperm count per ejaculate in our study was distinctly lower (1.86 × 106). However, loss of sperm due to adherence to the forceps and the copulatory plug cannot be ruled out. Separation of the ejaculate from the plug can be tricky due to the uneven surface of the plug. We did not determine the volumes of the viscous ejaculates to avoid any negative effects on sperm quality due to aspiration. The lower number of spermatozoa and the large variation between the collected ejaculates may further suggest that the influence of the separation procedure immediately after the onset of the ejaculatory shudder reduces the amount of spermatozoa released in the male’s ejaculate. Alternatively, the B6D2F1 hybrid males used in our study might a priori have a lower sperm count than wild-caught Mus m. domesticus.
Another possible factor influencing the amount of released sperm could be the successive use of the same female mating partner. Once the sperm collection method was mastered in the present experiment, only one or two receptive females were required as a mating partner for the males on each day of the study. However, it has been reported that the mating behavior of males appears to be influenced by whether the female has been previously mated. The mating behavior of subsequent males was characterized by significantly more intromissions before ejaculation and a significantly longer latency to ejaculation [31]. Therefore, we wondered whether in our investigation a previously mated female could have an effect on the mating behavior and ejaculated sperm number of the subsequent male. In our study, we could not distinguish between mounts with and without intromission when observing mating behavior. However, both the number of mounts and the latency to ejaculation slightly increased if the female had previously mated with another stud male. Although these differences were statistically not significant and do not call the practicality of the semen collection method into question, they are in line with findings from previous research and are potentially interesting for reproductive biology.
For efficient archiving of mouse strains, a high yield of motile spermatozoa per donor male is advantageous. Using the standard method of obtaining mouse sperm from the cauda epididymis, the mean number of spermatozoa obtained from BALB/c males was 18.4 × 106 [32], and 10.0 × 106 for wild-caught male offspring from only one cauda epididymis [33]. In routine practice, the sperm collected from both cauda epididymides of a donor male is usually divided into ten freezing straws, each containing more than 1 × 106 sperm in 10 µl of sperm suspension. Even taking into account the losses due to freezing, this is far more sperm per straw than is required for successful IVF [34]. The recommended concentration of motile sperm for IVF is 1 × 106 to 2.5 × 106 sperm/mL [35]. With the sperm retrieval method presented, more than one ejaculate would normally be required to safely archive a line, which is feasible over a short period of time. Due to the high variability in the number of spermatozoa of successfully collected ejaculates, as shown in Supplementary Table 1, it may be necessary to repeat the procedure with the same donor in order to safely archive a strain. However, sperm collection with the presented method can be repeated as long as the donor is reproductively active.
With an average number of 1.86 × 106 sperm, the presented method achieves a greater sperm yield than electroejaculation. When different electroejaculation devices were tested, the total number of sperm per ejaculate varied between 0.05 × 106 and 1.4 × 106. In addition to the high donor mortality rate of up to 22%, a significant reduction in the motility and velocity of the retrieved sperm is another disadvantage of this method [12]. At 0.4 × 106 to 0.7 × 106, the amount of sperm obtained by puncturing the cauda epididymis was also significantly lower than that obtained by the introduced separation method [17, 19]. The use of a very small diameter cannula to aspirate sperm has potentially also negative effects on the quality of sperm [18]. Although repeated sperm retrieval is also possible using electroejaculation or epididymal puncture, it is important to note that both procedures can only be performed under anesthesia. Therefore, the significant stress of the donor animals has to be taken into consideration when evaluating these methods.
Collection of ejaculates from the reproductive tract of a freshly mated female provides a large sperm yield and could be repeated if necessary. However, this method requires the killing of the female. Sperm retrieved from the reproductive tract of previously mated females has been successfully used in IVF after cryopreservation [14]. The sperm from the ejaculates used in the here presented analysis were not previously cryopreserved, but their excellent fertilization ability was confirmed with an average of 76% of two-cell embryos after IVF (Table 1). In addition, the proportion of motile sperm in the collected ejaculates from all test males was usually at least 80%, which is a very good basis for cryopreservation (Fig. 6).
Several studies indicate qualitative differences between ejaculated mouse sperm and sperm obtained directly from the epididymis, which are mainly explained by the influence of bioactive substances in the seminal plasma. For example, in mouse ejaculates retrieved from the uteri of previously mated females, a greatly increased nuclease activity was detected in both spermatozoa and seminal plasma, which was much less pronounced in epididymal spermatozoa and epididymal fluid. This difference could lead to an increased susceptibility of ejaculated sperm to DNA damage under cryopreservation conditions, thus compromising the integrity of archived sperm [36]. Sperm retrieved from the uterus after ejaculation also differ from epididymal sperm in their swimming pattern and, most importantly, are more efficient at penetrating the cumulus-oocyte complex in vitro [37].
Capacitation of the sperm is a prerequisite for fertilization. A fundamental process of mammalian sperm capacitation is the efflux of cholesterol and phospholipids from the sperm membrane. For the maturation of epididymal spermatozoa, the TYH preincubation medium contains polyvinyl alcohol as a substitute for bovine serum albumin and β-cyclodextrins, which acts as cholesterol and phospholipid acceptors to induce and support the capacitation process [24, 38].
In contrast to epididymal spermatozoa, ejaculated spermatozoa are coated with seminal plasma proteins. The secretion of the mouse seminal vesicle is the major component of the seminal plasma and contains several seminal vesicle proteins (SVS1-7) that not only influence the formation of the copulatory plug but also control sperm fertility [39]. Using SVS2 knockout males, it has been shown that this seminal vesicle protein suppresses premature ectopic activation of sperm maturation in the uterus, thus protecting the integrity and survival of sperm in the female reproductive tract. As a result, almost all sperm from wild-type males still have an intact acrosome at the isthmus to the oviduct, and the acrosome reaction as the completion of the maturation process essentially only takes place in the ampulla of the oviduct. In the absence of SVS2, sperm begin to die in the uterine cavity, and their in vivo fertility is drastically reduced. In contrast, during IVF, epididymal spermatozoa from SVS2−/− males did not differ from wild-type spermatozoa in terms of motility and in vitro fertilization rate [40].
Seminal vesicle excision in male mice resulted in a greatly reduced pregnancy rate, similar to mating with SVS2 knockout males, due to abnormal development of the pre-implantation embryonic stages. In addition, abnormalities in growth and other metabolic parameters were observed in the few offspring [41]. Therefore, seminal plasma factors appear not only to ensure the survival and fertility of ejaculated sperm, but also to influence the developmental conditions of the embryos during pregnancy and the phenotypic characteristics of the offspring.
Cryopreserved sperm is routinely used in IVF to revitalize archived mouse lines. Whether there is a qualitative difference between ejaculated and epididymal spermatozoa for this application cannot be deduced from the available test results. However, from the results discussed on the influence of seminal plasma on fertilization success, it can be concluded that ejaculated sperm would be preferable for artificial in vivo insemination.
In the present study, males and females of an F1 hybrid strain were used to exploit the heterosis effect on fertility and specifically on male libido. However, inbred and hybrid strains of laboratory mice can differ significantly in their sexual behavior. It has been shown that C57BL/6 J males, the most commonly used inbred strain for mouse genome modification, mate faster than BALB/c or DBA/2 J males [42, 43]. The introduced collection method of ejaculates must therefore be tested in practice for the strain or genetic background in question.
In summary, the separation of mating mice at the onset of the ejaculation reaction is a simple and quick-to-learn method for obtaining reasonable amounts of good-quality mouse sperm in vivo. Thus, the presented method can contribute to the 3Rs concept by reducing the number of animals required compared to the standard procedure using epididymal sperm from dissected cauda epididymides. More importantly, it contributes to refinement, as all other existing techniques for repeated sperm collection are invasive and stressful for the animals. Finally, this method can be used to collect sperm not only for IVF or for archiving lines, but also, for example, for studies that require repeated sperm samples from the same individual over time.


留言 (0)