
記住我



The considerable therapeutic potential of human multipotent mesenchymal stromal cells (MSCs) has generated markedly increasing interest in regenerative medicine for indications such as bone regeneration, cartilage repair, regeneration of other musculoskeletal tissues, central nervous system rebuilding, and so on.1 However, investigators report studies of MSC using different methods of isolation and expansion and different approaches to characterizing the cells.2 Thus it is increasingly difficult to compare and contrast study outcomes, which hinders progress in the field. To begin to address this issue, the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy proposes minimal criteria to define human MSC.2 First, MSC must be plastic-adherent when maintained in standard culture conditions. Second, MSC must express CD105, CD73, and CD90, and lack expression of CD45, CD34, CD14, or CD11b, CD79α, or CD19 and HLA-DR surface molecules. Third, MSC must differentiate into osteoblasts, adipocytes, and chondroblasts in vitro.2 While these criteria will probably require modification as new knowledge unfolds, we believe this minimal set of standard criteria will foster a more uniform characterization of MSC and facilitate the exchange of data among investigators.
MSCs are multipotent adult stem cells with significant regenerative potential and have garnered considerable interest in regenerative medicine and cell-based therapies.1 Despite their applicability, MSCs are usually derived from bone marrow or fat, both of which are invasive and the source of donors varies, resulting in variable quality and limited applicability of MSCs.3,4 Therefore, find a suitable donor or an alternative production method for MSCs to solve the bottleneck encountered in the current mass production of MSCs.
ESCs are derived from the inner cell mass of early-stage embryos and possess the ability to differentiate into all three germ layers, including ectoderm, endoderm, and mesoderm, making them a robust and versatile cell source for generating MSCs.3,4 The differentiation potential of ESCs has been extensively studied, and various protocols have been developed to guide their differentiation into MSCs.4 On the other hand, induced pluripotent stem cells (iPSCs) are derived from somatic cells through reprogramming, using specific transcription factors to induce pluripotency, resembling the characteristics of ESCs.5 The discovery of iPSCs by Yamanaka and colleagues in 2006 revolutionized the field of regenerative medicine, offering the potential for personalized cell therapies and disease modeling without the ethical concerns associated with ESC research.6–9
iPSCs and ESCs are two remarkable types of stem cells that possess the ability to differentiate into various cell types, including MSCs.10 Both iPSCs and ESCs offer the advantage of being able to generate an unlimited supply of MSCs, overcoming the limitations posed by donor availability in traditional MSC-based therapies.11 While both iPSCs and ESCs hold great promise for generating MSCs, it is essential to comprehensively compare the characteristics and functionalities of MSCs derived from these two distinct sources.1,12,13 In addition, considering the ethical issues of ESCs and the higher accessibility of iPSCs in translation medicine, it’s plausible that iMSCs may carry potential advantages over eMSCs in therapeutic applications. Collectively, understanding the similarities and differences between iPSC-derived MSCs (iMSCs) and ESC-derived MSCs (eMSCs) is crucial for determining their suitability for specific regenerative medicine applications. Additionally, a thorough comparative analysis will shed light on the potential advantages and limitations of each approach, informing the selection of the most appropriate source of MSCs for various therapeutic interventions.
Despite the widespread application of stem cells in various clinical disease models, where they can replace damaged tissues or promote cell regeneration, challenges persist in obtaining stem cells. The sources of donor cells lack uniformity, conditions for cell culture and differentiation are inconsistent, and there are risks associated with rejection reactions during transplantation. To overcome these issues regarding immune compatibility, iPSC technology has rapidly advanced to promote the matching of human leukocyte antigen (HLA) haplotype profile between the donors and recipients, providing an opportunity for allogeneic iPSC transplantations without the need for immune suppression. This evolution of iPSC technologies allowed the generation of HLA-homozygous iPSCs and other iPSC-derived lineages, leading to the expansion of their translational application. In our previous research, we successfully established highly immune-compatible iPSC cell lines (A33:03-B58:01-DRB1*03:01).14 In this study, we aimed to explore the differences between highly immune-compatible iPSC lines and ESCs and whether there are variations in their ability to differentiate into MSC cells. We aimed to provide a comprehensive comparison of iPSC-derived MSCs and ESC-derived MSCs. We examined their differentiation efficiency, cellular morphology, surface marker expression, and multilineage differentiation potential. By elucidating the distinct characteristics of MSCs derived from iPSCs and ESCs, we sought to contribute to the ongoing advancement of regenerative medicine and facilitated the development of more effective and tailored cell-based therapies.
2. METHODS 2.1. Establishment of human iPSCs 2.1.1. Derivation of human induced pluripotent stem cellsHuman iPSCs were derived from peripheral blood mononuclear cells (PBMCs) using the CytoTune-iPS 2.0 Sendai Reprogramming Kit. PBMCs were cultured in StemPro-34 medium with cytokines for 4 days, infected with Sendai virus, and then cultured in PBMC complete medium for 2 days. They were then transferred to MEF feeders and cultured in hESCs medium for 7 days.
Multiple colonies of iPSCs were generated and examined. Following the generation of HLA-homologous iPSCs (clone-1), we compared the ability for adipogenic and osteogenic differentiation among HLA-homologous iPSCs, control iPSCs (NTA, a previously established iPSC line from a healthy normal donor), H9 ESCs (a widely used ESC line), and S6 HSCs (an ESC line provided from Academia Sinica as a generous gift).
2.1.2. ESCs and iPSCs maintenance cultureESCs and iPSCs were cultured in StemFlex™ Medium Kit (ThermoFisher, Waltham, USA) on Geltrex™-coated dishes. Geltrex™ matrix was diluted with cold D-MEM/F-12 (1X) (ThermoFisher) and incubated in a humidified incubator at 37°C and 5% CO2 for at least 1 hour before use. Sub-culturing of iPSCs was performed using Versene solution (ThermoFisher).
2.2. Establishment of ESCs/iPSCs-derived mesenchymal stem cellsTo establish iPSC-derived mesenchymal stem cells, we followed the protocol described by Hynes et al15 with some modifications. Briefly, iPSCs were dissociated using Versene solution, and then reseeded onto a new Geltrex-coated T-25 flask for differentiation into mesenchymal stem cells in complete medium for 2 weeks, with medium replaced every 3 to 4 days. After the differentiation period, the heterogeneous cell population was passaged using TrypLE to obtain a single-cell suspension, which was then plated at a 1:3 ratio in a Geltrex-coated T-75 flask. Passaging was performed once cells reached 70% to 80% confluency, and cells were defined as passage 1 (P1) after the first passaging. From passage 2 onward, coating of the flask was not required.
2.2.1. Osteogenic differentiationOsteogenic differentiation was induced using StemPro™ Osteogenesis Differentiation Kit (Thermo Fisher Scientific, cat. no. A1007201) according to the manufacturer’s instructions. The medium was changed every 3 to 4 days for >21 days. Alizarin red staining (Sigma-Aldrich, St. Louis, USA 2003999) of calcium deposits within osteocytes generated by MSCs.
2.2.2. Semi-quantitative alizarin red stainingStained cultures were destained by a quantitative destaining procedure using 10% (w/v) cetylpyridinium chloride (CPC) (Sigma-Aldrich) in 10 mM sodium phosphate buffer, pH 7.0, for 15 min at room temperature on a vibrating platform shaker. The Alizarin Red S concentration was determined by absorbance measurement at 570 nm on a microplate reader using an Alizarin Red S standard curve in the same solution.
2.2.3. Adipogenic differentiationThe Adipogenic differentiation was performed using StemPro™ Adipogenesis Differentiation Kit (Thermo Fisher) and accorded to the manufacturer’s instructions. Culture cells in their respective medium for 7 to 14 days with every 3 to 4 days medium changes. The cells were then stained with filter Oil Red O solution prepared from Oil Red O solution (Sigma-Aldrich, O1391) for at least 2 hours at room temperature in the dark.
2.2.4. Semi-quantitative Oil Red O stainingOil Red O stain was eluted with 100% isopropanol, 500 µL per well for 10 minutes at room temperature on a vibrating platform shaker. 2 × 200 µL of eluate was transferred to a clear 96-well microtiter plate; a duplicate of wells on the microtiter plate was filled with 2 × 200 µL 2-propanol as background control to subtract the background signal. Absorption was measured at 510 nm on a microplate reader.
2.2.5. Chondrogenesis differentiationThe chondrogenic differentiation was performed using StemPro™ Chondrogenesis Differentiation Kit (Thermo Fisher) and accorded to the manufacturer’s instructions. Alcian Blue stain (Sigma-Aldrich, B8438) analysis: Alcian Blue staining of proteoglycans within chondrocytes generated by MSCs.
2.2.6. Flow cytometryMesenchymal stem cells were seeded in a 10-cm culture dish at a density of 2.5 × 103/cm2. Upon reaching 70% to 80% confluency, cells were collected using TrypLE Select Enzyme (Thermo Fisher, Cat. 12563011) and incubated with conjugated antibodies including CD14 (abcam, ab221678), CD19 (abcam, ab245235), CD34 (abcam, ab81289), CD45 (abcam, ab40763), HLA-DR (abcam, ab92511), TRA-1-81(Invitrogen, 17-8883-42), CD44 (abcam, ab254530), CD73 (abcam, ab313339), CD90 (abcam, ab307736), and CD105 (abcam, ab252345) in 3% Bovine serum albumin (BSA) in Phosphate buffered saline (PBS) for 35 minutes at room temperature in the dark. After washing with 10% FBS in PBS, cells were analyzed using FACSCanto II™ (BD, Franklin Lakes, NJ, USA) System and FlowJo™ software (BD, Franklin Lakes, NJ, USA).
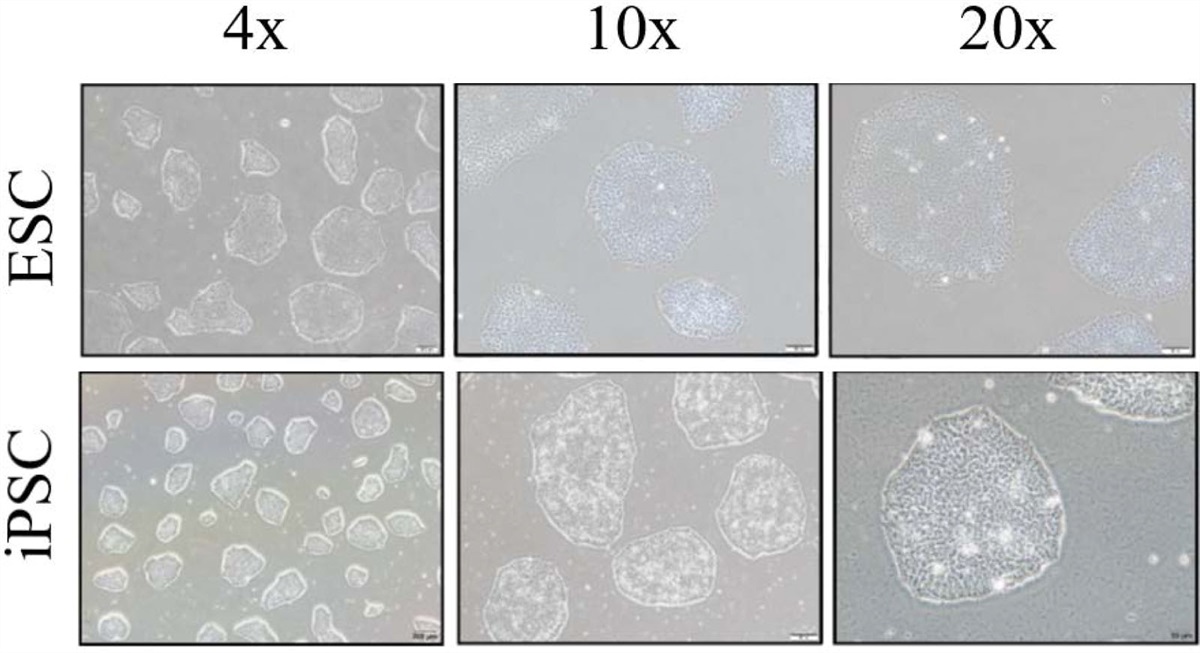
3. RESULTS 3.1. Acquisitions of iPSC and ESCsBefore the initiation of this study, we created a human iPSC line from the PBMCs obtained from donors carrying the HLA type (A33:03-B58:01-DRB1*03:01) as described previously.14 The reprogramming process involved infecting the PBMCs with Sendai virus, which expressed four Yamanaka factors, that is, OCT4, SOX2, KLF4, and c-MYC. We used the CytoTune-iPS 2.0 Sendai Reprogramming Kit from ThermoFisher A16518 to carry out this process. After the infection and cultivation, we generated the new iPSC line. To conduct a comparative study of the differentiation potential, we also enrolled an ESC line named S6 into this study. To validate the qualities of iPSCs and ESCs, we captured the morphology of both iPSCs and ESC (S6) under a light microscope. Both cell lines exhibited the typical features of pluripotent stem cells, including small and tightly packed cells, a high nucleus to cytoplasm ratio, compact colonies with distinct borders, and well-defined edges (Fig. 1).
 Fig. 1:
Fig. 1: Cell morphology of iPSCs and ESCs cell lines. iPSC line from the HLA-homozygous donors and ESCs cell (S6) both display a typical pluripotent stem cell’s morphology with small, tightly packed cells, high nucleus to cytoplasm ratio, compact colonies that have distinct borders, and well-defined edges. Scale bar = 200 μm (4×), 100 μm (10×), 50 μm (20×). ESCs = embryonic stem cells; HLA = human leukocyte antigen; iPSCs = induced pluripotent stem cell.
3.2. Derivation of MSCs from ESCs and iPSCsTo establish ESC/iPSC-derived MSCs, we followed the protocol described by Hynes et al15 with some modifications. Cell morphologies were significantly changed during the induction course of MSCs from both ESCs and iPSCs (Fig. 2A, B). Before the induction, ESCs and iPSCs exhibited an oval shape and were densely packed. However, after 14 days in the MSC complete medium, both ESCs and iPSCs began to expand, displaying a morphology more reminiscent of fibroblasts (as shown in Fig. 2A, B). Upon passaging onto gelatin-coated flasks, the differentiated ESCs and iPSCs adopted the typical morphology of MSCs. They featured elongated cell bodies and slender, elongated processes. Changes in cell morphology were also noted at various induction stages, with cells gradually adopting a fibroblast-like appearance as they progressed through the differentiation process. To differentially compare the MSCs from two pluripotent stem cell lines, ESC-derived MSCs and iPSC-derived MSCs were designated as eMSCs and iMSCs, respectively. This nomenclature will be used consistently throughout the study.
 Fig. 2:
Fig. 2: The morphological changes during the differentiation from (A) ESC (S6) to eMSCs, and (B) HLA-homozygous iPSC to iMSC. (I) Undifferentiated ESC colonies before induction. (II) ESCs cultured in MSC complete medium for 14 d. (III) Differentiated ESCs passaged onto gelatin-coated flasks and grown in MSC complete medium for 14 d. (IV) Cell morphology alterations at different induction stages, including passages 2 and 8. (V) Enlarged views of regions are indicated by white boxes. Scale bar = 200 µm. ESCs = embryonic stem cells; HLA = human leukocyte antigen; iPSCs = induced pluripotent stem cell; MSC = human leukocyte antigen.
3.3. Comparative analysis of cell surface markers between eMSCs and iMSCsTo compare cell surface markers between eMSCs and iMSCs, we conducted a flow cytometry analysis. The assessment was based on the International Society for Cell & Gene Therapy (ISCT) criteria, where more than 95% of the MSCs were expected to express the markers CD73, CD90, and CD105.2 Additionally, the analysis aimed to ensure that <2% of the cells expressing CD14, CD19, CD34, CD45, and HLA-DR.2 Additionally, to provide a more comprehensive analysis, we also performed flow cytometry to estimate the expression of CD44 as another optional MSC marker and TRA-1-81 as a pluripotency marker in the analysis.
Flow cytometric analysis was performed to assess the expression of cell surface markers in ESCs and iPSCs before and after differentiation to MSCs (Figs. 3 and 4). In addition to the changes in cell morphology, the alterations in cell surface marker expression were also observed during the induction of ESCs/iPSCs to MSCs, and these differences are visually represented in the statistical charts (Figs. 3B and 4B). This graphical presentation allows for a direct comparison of the changes in surface marker expression when ESCs or iPSCs transitioned into MSCs. We assessed the expression profiles of 10 distinct markers (Figs. 3B and 4B). One noteworthy observation was the prominent presence of TRA-1-81 in parental ESCs and iPSCs, which exhibited a significant reduction following differentiation into eMSCs or iMSCs. This reduction is of particular significance as ESCs and iPSCs possess pluripotent characteristics and the potential to form teratomas. The decline in TRA-1-81 expression highlights the safety aspect of our established MSC lines for translational applications (Figs. 3 and 4). Furthermore, our flow cytometry analysis demonstrated that <2% of the derived cell population expressed hematopoietic markers, specifically CD14, CD19, CD34, and CD45. In contrast, over 95% of the derived cell population exhibited positive expression of essential MSC markers, including CD44, CD73, CD90, and CD105. Of equal importance, more than 98% of the derived cell population lacked HLA-DR expression (Figs. 3 and 4).
 Fig. 3:
Fig. 3: Flow cytometry analysis of ESC(S6) and eMSCs. A, Flow cytometry analysis was conducted to assess the expression of cell surface markers before and after differentiation. The blue histograms represent the isotype controls, while the red histograms represent the expression of individual markers evaluated in both ESC and eMSCs. B, eMSCs showed significantly higher expression of MSC markers in comparison with the parental ESC (S6). Results are mean ± SD, triplicated; *p ≤ 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. ESC = embryonic stem cell; MSCs = mesenchymal stem cells.
 Fig. 4:
Fig. 4: Flow cytometry analysis of HLA-homozygous iPSCs and iMSCs. A, Flow cytometry analysis was conducted to assess the expression of cell surface markers before and after differentiation. The blue histograms represent the isotype controls, while the red histograms represent the expression of individual markers evaluated in both iPSCs and iMSCs. B, iMSCs showed significantly higher expression of MSC markers in comparison with the parental iPSCs. Results are mean ± SD, triplicated; *p ≤ 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. HLA = human leukocyte antigen; iPSCs = induced pluripotent stem cells; MSCs = mesenchymal stem cells.
These findings collectively affirmed the MSC phenotypes of our derived cells, with a robust expression of characteristic MSC markers and the absence of hematopoietic markers, further emphasizing their potential suitability for regenerative medicine applications. The marked decrease in pluripotency marker TRA-1-81 underscores the safety and reliability of these cells for translational research and therapeutic purposes.
3.4. Comparative tri-lineage differentiation potential of iMSCs and eMSCsWe further explored and compared the differentiation potential of both eMSCs and iMSCs through three distinct assays. Chondrogenic differentiation was induced by subjecting these MSCs to a chondrogenic differentiation medium for 14 days. This led to the formation of chondrocyte pellets exhibiting the characteristic three-dimensional (3D) round morphology of chondrocytes (Fig. 5). Positive Alcian Blue staining validated the successful chondrogenesis from both eMSCs and iMSCs (Fig. 5).
 Fig. 5:
Fig. 5: Chondrogenesis of iMSCs and eMSCs. A, Representative images of chondrogenic capacity result of iMSCs were shown. Formed chondrocyte pellets shown by staining with Alcian Blue solution. Scale bar = 100 µm. B, Representative images of chondrogenic capacity result of eMSCs were shown. Formed chondrocyte pellets shown by staining with Alcian Blue solution. Scale bar = 100 µm. MSCs = mesenchymal stem cells.
For adipogenic differentiation, a specialized adipogenic differentiation medium was used. Lipid droplets emerged within the induced cells and were positively stained with Oil Red O solution (Fig. 6). Quantitative analysis of the extracted lipid droplets indicated a notably higher lipid content in adipocytes derived from both eMSCs and iMSCs compared to the control undifferentiated MSCs (Fig. 6).
 Fig. 6:
Fig. 6: Adipogenesis of eMSCs and iMSCs. A, Representative images of adipogenesis of eMSC (MSCs differentiated from H9 and S6 ESCs) and iMSCs (MSCs differentiated from control iPSCs [NTA] and HLA-homologous iPSCs). Morphologies of formed lipid droplets are shown by staining with Oil Red O solution, indicated by black arrows. Scale bar = 50 µm. B, Quantification of lipid elaborated by extraction of Oil Red O stain and measuring absorbance at O.D. 510 nm, n = 3 wells for each group. Results are mean ± SD from three independent experiments; *p ≤ 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. HLA = human leukocyte antigen; iPSCs = induced pluripotent stem cells; O.D. = optical density.
To assess the osteogenic potential, eMSCs and iMSCs were cultured in osteogenic differentiation medium for over 21 days. Alizarin Red S staining revealed bright orange-red deposits in the cells receiving osteogenic stimuli, indicating the successful formation of osteocytes from both eMSCs and iMSCs (Fig. 7). Conversely, the control undifferentiated MSCs exhibited no such deposits, emphasizing the successful osteogenic differentiation from both eMSCs and iMSCs.
 Fig. 7:
Fig. 7: Osteogenesis of eMSCs and iMSCs. A, Representative images of osteogenic of eMSC (MSCs differentiated from H9 and S6 ESCs) and iMSCs (MSCs differentiated from control iPSCs [NTA] and HLA-homologous iPSCs). Morphologies of mineral deposit are shown by staining with Alizarin Red S solution, indicated by white arrows. Scale bar = 100 µm. B, Representative images showing the osteogenic potential of eMSCs. Morphologies of mineral deposit are shown by staining with Alizarin Red S solution, indicated by white arrows. Scale bar = 100 µm. MSCs = mesenchymal stem cells. C, Quantification of mineral deposit elaborated by extraction of Alizarin Red S stain and measuring absorbance at O.D. 570 nm, n = 3 wells for each group. Results are mean ± SD from three independent experiments; *p ≤ 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. HLA = human leukocyte antigen; iPSCs = induced pluripotent stem cells; O.D. = optical density.
Overall, the differentiation potential of eMSCs and iMSCs into chondrocytes, adipocytes, and osteocytes were equivalent. These results collectively underscored the versatile differentiation potential of both eMSCs and iMSCs, making them promising candidates for various therapeutic applications.
4. DISCUSSIONIn this study, we successfully induced both ESCs and iPSCs generated from the donors with HLA type (A33:03-B58:01-DRB1*03:01) to differentiate into corresponding MSCs. The ability to generate MSCs from both ESCs and iPSCs offers a renewable and abundant source of these multipotent cells, overcoming the limitations posed by donor availability in traditional MSC-based therapies.16 This accomplishment not only expands the application spectrum of these stem cells but also prompts a closer examination of the differentiation methods used. It is intriguing to consider the differentiation methods used in generating MSCs from ESCs and iPSCs in the context of our findings.15,17–21 The development of methods that mimic natural embryonic developmental processes has been pivotal in ensuring the acquisition of predictable and functional MSCs. Notably, recent studies have proposed the induction of mesoderm as an intermediate step in MSC differentiation. This is consistent with the developmental origins of MSCs, particularly from the lateral plate mesoderm (LPM)22,23 and neural crest cells (NCCs).24–28 However, the present study did not directly explore the mesodermal induction step; our results align with the anticipated outcomes of this approach. The observed shifts in morphology and surface marker expression from pluripotent to mesenchymal characteristics reflect a comprehensive cellular transformation reminiscent of natural lineage commitment.
Our results demonstrated that the differentiated MSCs from both ESCs and iPSCs expressed high levels of key MSC markers, including CD73, CD90, and CD105. These findings indicate that the induced MSCs possess the potential to differentiate into various cell types, such as bone, cartilage, and fat, making them promising candidates for tissue repair and regeneration.1 The multilineage differentiation potential of the induced MSCs further highlights their suitability for therapeutic applications in treating various diseases and injuries.29,30 Notably, we observed that the differentiated MSCs from both ESCs and iPSCs expressed low levels of hematopoietic markers, such as CD14, CD19, CD34, and CD45.31 This finding suggests that the induced MSCs have a low potential to differentiate into hematopoietic lineage. This characteristic is advantageous for their application in regenerative medicine, as the distinct marker expression profiles in MSCs compared to hematopoietic stem cells indicate fundamental differences in their differentiation pathways.32 Our results confirm that both eMSCs and iMSCs express cell surface markers consistent with the MSC lineage, distinctly separate from those of hematopoietic stem cells. This alignment with the MSC marker profile enhances the safety and potential therapeutic utility of these induced MSCs. Furthermore, to assess the functional differentiation capacity of MSCs from both origins, we conducted comprehensive assays.33 Chondrogenic differentiation resulted in the formation of chondrocyte pellets displaying the characteristic 3D round morphology of chondrogenesis. Positive Alcian Blue staining further confirmed the successful chondrogenesis of eMSCs and iMSCs. In adipogenic differentiation assays, the appearance of lipid droplets within induced cells indicated the initiation of adipocyte differentiation.34 Quantitative analysis substantiated higher lipid content in adipocytes from both eMSCs and iMSCs, affirming their adipogenic differentiation potential. In the osteogenic differentiation assay, the presence of vivid orange-red deposits in the induction group signified successful osteocyte formation.35 The absence of such deposits in the control group highlighted the effective osteogenic differentiation capacity of both eMSCs and iMSCs. Despite the promising results, this study has certain limitations that warrant consideration. The study was conducted with a small number of cells, and further investigations using larger populations of cells are needed to validate and extend our findings. Additionally, while our study focused on the differentiation of MSCs, future research should explore the functional properties and potential therapeutic effects of these induced MSCs in relevant disease models.
Moreover, despite the equivalent performance of eMSCs and iMSCs in functional differentiation into adipocytes, chondrocytes, and osteocytes, it is essential to consider the potential advantages of using iMSCs over eMSCs in various therapeutic applications. First, it is difficult to avoid the ethical issues regarding ESCs in the use of eMSCs. On the contrary, the ability to generate iPSCs and corresponding iMSCs from readily accessible adult tissues offers a significant advantage in terms of availability and ease of procurement, making them an attractive and potentially inexhaustible source for regenerative medicine. Recent studies have indicated that iMSCs exhibit greater proliferation potential and can be derived using various efficient methods.36,37 These characteristics, combined with their promising preclinical and clinical therapeutic potential, position iMSCs as a compelling alternative to traditional eMSCs.36
In conclusion, our study demonstrates the successful and comparable induction of MSCs from both ESCs and iPSCs, providing a renewable and abundant source of these multipotent cells for regenerative medicine applications. The similar characteristics observed in the induced MSCs from ESCs and iPSCs indicate their potential equivalence in therapeutic efficacy. Remarkably, unlike ESCs that carry ethical issues, iPSCs from donors matching HLA haplotype profiles eliminate the issue of long-term immunosuppressant use. After differentiation into MSCs, HLA-matched donor-derived iMSCs that are safer for patients contribute to the progress of stem cell-based therapies, opening up new avenues for personalized and effective treatments for various diseases and injuries. Future research endeavors can further explore the therapeutic potential of these induced MSCs and enhance our capabilities in harnessing the regenerative power of stem cells.
ACKNOWLEDGMENTSGrateful acknowledgment is extended for the technical support provided by Hsiao-Yun Tai, Yi-Ching Tsai, and Pei-Yu Weng from the Department of Medical Research, Taipei Veterans General Hospital.
REFERENCES 1. Han Y, Li X, Zhang Y, Han Y, Chang F, Ding J. Mesenchymal stem cells for regenerative medicine. Cells. 2019;8:886. 2. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7. 3. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–7. 4. Hawkins KE, Corcelli M, Dowding K, Ranzoni AM, Vlahova F, Hau KL, et al. Embryonic stem cell-derived mesenchymal stem cells (MSCs) have a superior neuroprotective capacity over fetal MSCs in the hypoxic-ischemic mouse brain. Stem Cells Transl Med. 2018;7:439–49. 5. Ye L, Swingen C, Zhang J. Induced pluripotent stem cells and their potential for basic and clinical sciences. Curr Cardiol Rev. 2013;9:63–72. 6. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. 7. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–72. 8. Lee CY, Huang CH, Rastegari E, Rengganaten V, Liu PC, Tsai PH, et al. Tumor necrosis factor-alpha exacerbates viral entry in SARS-CoV2-infected iPSC-derived cardiomyocytes. Int J Mol Sci. 2021;22:9869. 9. Chien CS, Chien Y, Lin YY, Tsai PH, Chou SJ, Aliaksandr AY, et al. Dual DNA transfection using 1,6-Hexanedithiol-Conjugated Maleimide-Functionalized PU-PEI(600) for gene correction in a patient iPSC-derived Fabry cardiomyopathy model. Front Cell Dev Biol. 2021;9:634190. 10. Zomer HD, Vidane AS, Gonçalves NN, Ambrósio CE. Mesenchymal and induced pluripotent stem cells: general insights and clinical perspectives. Stem Cells Cloning. 2015;8:125–34. 11. Zhao C, Ikeya M. Generation and applications of induced pluripotent stem cell-derived mesenchymal stem cells. Stem Cells Int. 2018;2018:1–8. 12. Mazini L, Rochette L, Amine M, Malka G. Regenerative capacity of adipose derived stem cells (ADSCs), comparison with mesenchymal stem cells (MSCs). Int J Mol Sci. 2019;20:2523. 13. Ju Y, Yi L, Li C, Wang T, Zhang W, Chai W, et al. Comparison of biological characteristics of human adipose- and umbilical cord- derived mesenchymal stem cells and their effects on delaying the progression of osteoarthritis in a rat model. Acta Histochem. 2022;124:151911. 14. Liu DH, Tseng HC, Lee MS, Chiou GY, Wang CT, Lin YY, et al. Overcoming the challenges of scalable iPSC generation in translation medicine. J Chin Med Assoc. 2024;87:163–70. 15. Hynes K, Menicanin D, Gronthos S, Bartold MP. Differentiation of iPSC to mesenchymal stem-like cells and their characterization. Methods Mol Biol. 2016;1357:353–74. 16. Luzzani CD, Miriuka SG. Pluripotent stem cells as a robust source of mesenchymal stem cells. Stem Cell Rev Rep. 2017;13:68–78. 17. Olivier EN, Rybicki AC, Bouhassira EE. Differentiation of human embryonic stem cells into bipotent mesenchymal stem cells. Stem Cells. 2006;24:1914–22. 18. Hwang NS, Varghese S, Lee HJ, Zhang Z, Ye Z, Bae J, et al. In vivo commitment and functional tissue regeneration using human embryonic stem cell-derived mesenchymal cells. Proc Natl Acad Sci U S A. 2008;105:20641–6. 19. Karlsson C, Emanuelsson K, Wessberg F, Kajic K, Axell MZ, Axell PS, et al. Human embryonic stem cell-derived mesenchymal progenitors—potential in regenerative medicine. Stem Cell Res. 2009;3:39–50. 20. Boyd NL, Robbins KR, Dhara SK, West FD, Stice SL. Human embryonic stem cell-derived mesoderm-like epithelium transitions to mesenchymal progenitor cells. Tissue Eng Part A. 2009;15:1897–907. 21. Lian Q, Zhang Y, Liang X, Gao F, Tse HF. Directed differentiation of human-induced pluripotent stem cells to mesenchymal stem cells. Methods Mol Biol. 2016;1416:289–98. 22. Mahmood A, Harkness L, Schrøder HD, Abdallah BM, Kassem M. Enhanced differentiation of human embryonic stem cells to mesenchymal progenitors by inhibition of TGF-beta/activin/nodal signaling using SB-431542. J Bone Miner Res. 2010;25:1216–33. 23. Sánchez L, Gutierrez-Aranda I, Ligero G, Rubio R, Muñoz-López M, García-Pérez JL, et al. Enrichment of human ESC-derived multipotent mesenchymal stem cells with immunosuppressive and anti-inflammatory properties capable to protect against experimental inflammatory bowel disease. Stem Cells. 2011;29:251–62. 24. Takashima Y, Era T, Nakao K, Kondo S, Kasuga M, Smith AG, et al. Neuroepithelial cells supply an initial transient wave of MSC differentiation. Cell. 2007;129:1377–88. 25. Morikawa S, Mabuchi Y, Niibe K, Suzuki S, Nagoshi N, Sunabori T, et al. Development of mesenchymal stem cells partially originate from the neural crest. Biochem Biophys Res Commun. 2009;379:1114–9. 26. Fukuta M, Nakai Y, Kirino K, Nakagawa M, Sekiguchi K, Nagata S, et al. Derivation of mesenchymal stromal cells from pluripotent stem cells through a neural crest lineage using small molecule compounds with defined media. PLoS One. 2014;9:e112291. 27. Menendez L, Kulik MJ, Page AT, Park SS, Lauderdale JD, Cunningham ML, et al. Directed differentiation of human pluripotent cells to neural crest stem cells. Nat Protoc. 2013;8:203–12. 28. Chambers SM, Fasano CA, Papapetrou EP, Tomishima M, Sadelain M, Studer L. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat Biotechnol. 2009;27:275–80. 29. Mishra VK, Shih HH, Parveen F, Lenzen D, Ito E, Chan TF, et al. Identifying the therapeutic significance of mesenchymal stem cells. Cells. 2020;9:1145. 30. Thanaskody K, Jusop AS, Tye GJ, Wan Kamarul Zaman WS, Dass SA, Nordin F. MSCs vs. iPSCs: potential in therapeutic applications. Front Cell Dev Biol. 2022;10:1005926. 31. Lachmann N, Ackermann M, Frenzel E, Liebhaber S, Brennig S, Happle C, et al. Large-scale hematopoietic differentiation of human induced pluripotent stem cells provides granulocytes or macrophages for cell replacement therapies. Stem Cell Rep. 2015;4:282–96. 32. Battiwalla M, Hematti P. Mesenchymal stem cells in hematopoietic stem cell transplantation. Cytotherapy. 2009;11:503–15. 33. Bianco P, Robey PG, Simmons PJ. Mesenchymal stem cells: revisiting history, concepts, and assays. Cell Stem Cell. 2008;2:313–9. 34. Safwani WK, Makpol S, Sathapan S, Chua K. Impact of adipogenic differentiation on stemness and osteogenic gene expression in extensive culture of human adipose-derived stem cells. Arch Med Sci. 2014;10:597–606. 35. Phillips MD, Kuznetsov SA, Cherman N, Park K, Chen KG, McClendon BN, et al. Directed differentiation of human induced pluripotent stem cells toward bone and cartilage: in vitro versus in vivo assays. Stem Cells Transl Med. 2014;3:867–78. 36. Arakawa M, Sakamoto Y, Miyagawa Y, Nito C, Takahashi S, Nitahara-Kasahara Y, et al. iPSC-derived mesenchymal stem cells attenuate cerebral ischemia-reperfusion injury by inhibiting inflammatory signaling and oxidative stress. Mol Ther Methods Clin Dev. 2023;30:333–49. 37. Sabapathy V, Kumar S. hiPSC-derived iMSCs: NextGen MSCs as an advanced therapeutically active cell resource for regenerative medicine. J Cell Mol Med. 2016;20:1571–88.
留言 (0)