
記住我

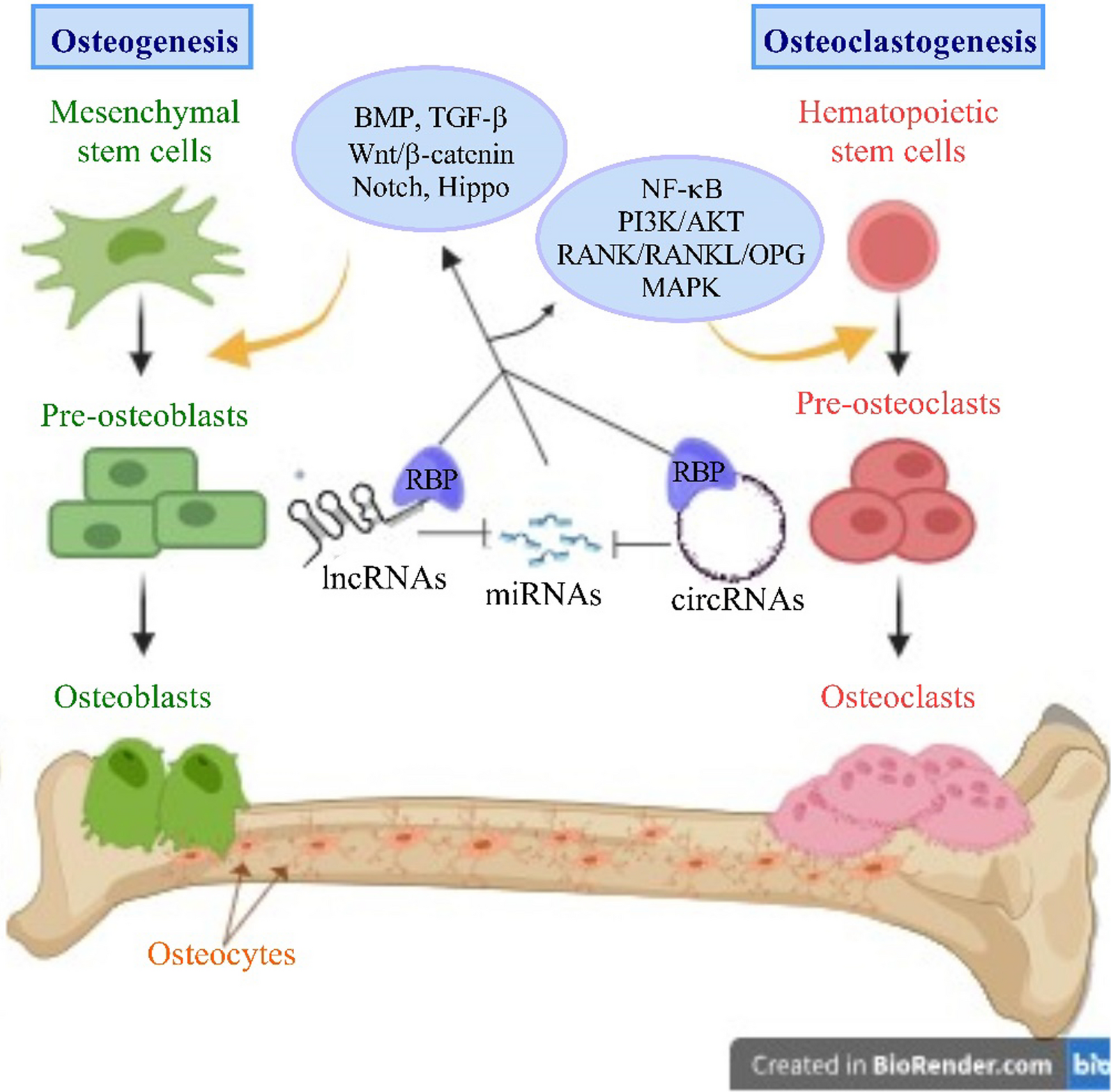

Osteogenesis is the differentiation of committed mesenchymal stem cells (MSCs) into mature, active osteoblasts through several stages of precursors. Osteogenesis is achieved through interrelated cascades including hippo, Wnt, notch, and bone morphogenetic protein (BMP). These axes modulate diverse transcription factors, such as runt-related transcription factor 2 (RUNX2). RUNX2 participates in triggering the dedication of MSCs to the osteogenic lineage and functions upstream from the other transcription factors specific to osteoblasts, such as osterix (Osx) and distal-less homeobox 5 (Dlx5) [16]. The Wnt/β-catenin pathway plays a potential role in the stabilization of β‐catenin that sequentially migrates to the nucleus and controls the expression of genes related to osteoblastogenesis [17]. Dickkopf-1 (DKK1) is mainly expressed by osteoblasts and bone marrow stromal cells (BMSCs) and antagonizes the Wnt pathway.
Another signaling pathway is BMP, which is one of the members of the transforming growth factor (TGF)-β superfamily and plays a crucial role in regenerating osteogenic differentiation in osteoblasts. Briefly, BMP receptors bind with BMP ligands, resulting in phosphorylation of Smad1/5/8, which then attaches to the co-Smad (Smad4) to form a complex. The resultant complex moves to the nucleus, where it regulates the genes that affect osteoblast development in conjunction with co-activators and corepressors [18].
The Hippo signaling cascade modulates osteoblastic activity and thereby participates in bone development and maintenance. Sometimes, the Hippo pathway is turned off by inactivating (dephosphorylating) mammalian STE20-like protein kinases 1 and 2 (MST1/2) and large tumor suppressors 1 and 2 (LATS1/2). This results in the activation of the yes-associated protein (YAP) and the transcriptional coactivator with the PDZ-binding motif (TAZ), allowing them to translocate into the nucleus. Subsequently, YAP and TAZ act as transcriptional co-activators, interacting with transcription factors to regulate the expression of target genes related to osteoblast cell proliferation, differentiation, and survival [19].
Notch signaling, a highly preserved intercellular pathway, governs cell differentiation and proliferation as well as determines cell fate via the activation of cell surface receptors (Notch 1-4) and their delta-like ligands (DLL) 1, 3, 4, and Jagged 1, 2. This pathway promotes the expression of downstream target genes involved in osteogenesis regulation [20].
LncRNAs involved in osteogenesis regulationLncRNAs that promote osteogenesisBMSCs have a high osteogenic capability, which is considered an effective practical strategy for hindering bone loss. Previous reports elucidated the involvement of the lncRNAs in BMSC proliferation and differentiation. Herein, we review the expression of potential lncRNAs in osteoporosis and their significant roles in either promoting or inhibiting osteogenic differentiation, as shown in Table 1 and Fig. 2, respectively.
Table 1 The expression of lncRNAs-related osteogenesis in osteoporosisFig. 2
Role and mechanistic insights of lncRNAs in osteogenesis. All shapes shaded in green denote biomarkers increased during osteogenesis. All shapes shaded in red denote biomarkers decreased during osteogenesis
LncRNA homeobox D gene cluster antisense growth-associated long noncoding RNA (HAGLR) exhibited a significant decrease in the peripheral blood of patients with postmenopausal osteoporosis (PMOP) compared to healthy controls. Conversely, its expression increased during BMSC osteogenic differentiation. In OVX mice, the overexpression of HAGLR alleviated PMOP by upregulating homeobox protein A10 (Hoxa10) expression through inhibition of miR-182-5p [21]. Indeed, Hoxa10, a crucial transcription factor in the osteogenic process, plays a pivotal role in regulating the production of osteoblasts [22]. In parallel, the lncRNA RAD51-Antisense RNA 1 (RAD51-AS1) was remarkably downregulated in human bone marrow stem cells (hBMSCs) obtained from osteoporotic patients compared to those from controls. It was observed that RAD51-AS1 knockdown inhibited the proliferation and differentiation of hBMSCs and boosted their apoptosis. Mechanistically, RAD51-AS1 was determined to stimulate the osteogenic differentiation of hBMSCs by interacting with Y-Box Binding Protein 1 (YBX1) to generate mRNA-protein complexes with SMAD7 and Smad ubiquitin regulatory factor 2 (SMURF2), thus hampering their translation. Additionally, it could transcriptionally upregulate proliferating cell nuclear antigen (PCNA) and SIVA1 by binding to their promoter regions, resulting in activation of the TGF-β signaling pathway [23]. LncRNA KCNQ1OT1 also showed a significant decrease in PMOP patients compared to control group. Its overexpression enhanced the osteogenic differentiation of MC3T3-E1 cells and alleviated osteoporosis in OVX mice through upregulation of mTOR via sponging miR-421-3p [24].
Besides, GATA-binding protein 4 (GATA4) is crucial in modulating osteoblastic differentiation, bone remodeling, and mineralization [25]. Huang et al. linked GATA4 with lncRNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) in BMSCs during osteogenesis [26]. The study found that GATA4 overexpression promoted BMSC osteoblastic differentiation through upregulating lncRNA MALAT1. Notably, the expression of both GATA4 and lncRNA MALAT1 was reduced in the bone tissues of both PMOP patients and ovariectomized (OVX) mice. It was confirmed that GATA4 activated the transcription of MALAT1, which in turn formed an RBP complex with K-homology splicing regulatory protein (KHSRP) to decay the stability of neuronal precursor cell-expressed developmentally downregulated 4 (NEDD4) mRNA, the one that is responsible for the promotion of RUNX1 degradation by ubiquitination [27]. In sum, GATA4-mediated lncRNA MALAT1 stimulated osteogenic differentiation of BMSCs via inhibiting RUNX1 degradation through modulation of the KHSRP/NEDD4 axis, ultimately driving bone formation in PMOP. Furthermore, another study has also investigated the role and molecular mechanism of MALAT1 in osteoporosis. Interestingly, several phytochemicals have been found to be effective therapeutics for osteoporosis. Among them is quercetin, which has a promoting effect on BMSC osteogenesis [28]. Feng et al. documented that quercetin treatment could attenuate tumor necrosis factor alpha (TNFα)-impaired BMSCs osteogenesis via upregulating MALAT1 expression [29]. In OVX mice, serum MALAT1 level was markedly decreased, while quercetin treatment restored its level to some extent. Additionally, MALAT1 was involved in the modulating effect of quercetin on TNFα-impaired BMSC osteogenesis. Its knockdown abolished the rescuing effect of quercetin; stimulation of β-catenin expression and mitigation of NF-κB p65 translocation. This, in turn, compromised quercetin's osteogenic ability and facilitated the progression of osteoporosis. In the same context, Paired-Related Homeobox Protein 2 (Prrx2) is a transcription factor that belongs to the paired-related homeobox protein family and has been found to be involved in the regulation of lncRNA expression. Li et al. reported that myoblast-derived exosomal Prrx2 directly attached to the MIR22HG promoter and enhanced its expression [30]. Relevantly, high levels of MIR22HG alleviated OVX-induced bone loss by accelerating BMSC osteogenic differentiation through inhibition of miR-128, which in turn stimulated YAP1 expression and thus activated the Hippo pathway.
Other factors have been implicated in the regulation of lncRNAs during osteogenesis, impacting the development of osteoporosis. Dexamethasone (DEX) is a synthetic glucocorticoid that has been reported to regulate MSC osteogenic lineages. LncRNA TERC is among the lncRNAs identified to alleviate DEX-induced osteoporosis. It was observed that TERC overexpression mitigated the inhibitory effect of DEX on BMSC osteogenic differentiation by increasing the level of the enhancer of zeste homolog 2 (EZH2) protein. This elevation, in turn, facilitated the histone modification of DKK1, thereby activating the Wnt signaling pathway. Thus, TERC overexpression by lessening DEX-induced osteoporosis could provide an additional therapeutic strategy for glucocorticoid-induced osteoporosis [31]. Also, Autophagy is a mechanism beneficial for cell survival and growth [32], known to maintain bone remodeling by removing reactive oxygen species (ROS) [33]. Xue et al. have found that lncRNA Small nucleolar RNA host gene 14 (SNHG14) overexpression activated autophagy in BMSCs as well as raised bone mineral density (BMD) and bone trabecular number in OVX mice [34]. Mechanically, an elevated level of SNHG14 stimulated myocyte enhancer factor 2C (Mef2c)-mediated autophagy activation through inhibition of miR-493-5p, predisposing to enhancement of the osteogenic ability of BMSCs. Thus, SNHG14 could alleviate osteoporosis progression via activation of autophagy by regulating the miR-493-5p/Mef2c axis.
LncRNAs that inhibit osteoblastogenesisCertain lncRNAs have been identified as inhibitors of osteogenesis, which impede the process of bone formation and trigger osteoporosis development. For example, LINC00205 was upregulated in bone tissue samples of osteoporotic patients with or without a spinal fracture, while it was downregulated during the osteogenic differentiation of human mesenchymal stem cells (hMSCs). Precisely, LINC00205 inhibited osteogenic differentiation by targeting miR-26b-5p and enhancing lysine (K)-specific methyltransferase 2C (KMT2C) expression. This latter contributed to a decrease in the expression of osteogenic-related genes such as RUNX2, alkaline phosphatase (ALP), and osteocalcin (OCN), resulting in the progression of osteoporosis [35]. Likewise, LINC01234 was highly expressed in the plasma of patients with osteoporosis while exhibiting a gradual decrease during osteogenic differentiation of hMSCs. In this study, it was observed that LINC01234 impeded hMSC osteogenic differentiation by augmenting aldehyde oxidase 1 (AOX1) via regression of miR-513a-5p. Notably, the AOX1 gene encodes the cytosolic enzyme that catalyzes the formation of superoxide [36]. Thus, targeting LINC01234 could promote osteogenesis and hinder osteoporosis development [37].
LncRNA PCBP1-AS1 was highly expressed in osteoporotic tissues and decreased during the development of hBMSCs into osteoblasts. The knockdown of PCBP1-AS1 promoted the osteogenic differentiation capacity of hMSCs, while its overexpression exhibited an opposite effect. Mechanistically, PCBP1-AS1 targeted Pak family member p21-activated kinase 2 (PAK2) by inhibiting miR-126-5p [38]. Remarkably, PAK2 is required for various cellular activities, such as cytoskeletal remodeling and chromatin modulation [39]. Additionally, lncRNA KCNMA1-AS1 was elevated in the osteoporotic subjects. Its suppression upregulated miR-1303, which resulted in the downregulation of cochlin (COCH), a known suppressor of embryonic stem cell differentiation. This led to the promotion of osteogenic differentiation in hMSCs and the mitigation of the progression of osteoporosis [40].
The intricate interplay between lncRNAs and apoptosis reveals a novel aspect of the pathogenesis of osteoporosis. Pan et al. reported that lncRNA SNHG1 could regulate osteogenic differentiation by influencing pyroptosis, an inflammatory type of apoptosis triggered by inflammasomes and caspase-1 [41]. SNHG1 was observed to be elevated in the serum of osteoporotic patients. Its overexpression could suppress BMSC osteogenic differentiation through interaction with HMGB1, enhancing its expression. This, in turn, activated pyroptosis-associated factors (caspase-1 p20 and gasdermin D-N (GSDMD)-N) and the production of inflammatory interleukins (IL-1β and IL-18), leading to the development of osteoporosis. In tandem, interferon regulatory factor-1 (IRF-1) also participated in controlling cellular apoptosis during osteogenic differentiation through the regulation of lncRNA X-inactive-specific transcript (XIST). Elevated levels of lncRNA XIST were observed in OVX mice, and its knockdown not only alleviated osteoporosis symptoms but also promoted osteogenic differentiation in hBMSCs. Simultaneously, IRF-1 accelerated osteogenic differentiation by repressing the transcription of XIST, which in turn upregulated miR-450b and subsequently decreased F-box and WD repeat domain-containing 7 (FBXW7) expression [42]. FBXW7 is regarded as an efficient tumor suppressor due to its role in cellular apoptosis and has also been observed to be involved in the osteogenic differentiation of BMSCs [43, 44].
CircRNAs involved in osteogenesis regulationCircRNAs that promote osteogenesisSeveral studies, conducted in BMSC-induced osteogenic differentiation, focused on RUNX2 regulation through various circRNA/miRNA axes. For instance, circ-3626, an exonic circRNA that arises from the STAG1 gene, was significantly increased during osteogenic differentiation of BMSCs, whereas it was decreased in bone tissues derived from osteoporotic patients and BMSCs of aged mice. Moreover, its overexpression dramatically accelerated the osteogenic capability of BMSCs. Circ-3626 increased RUNX2 regulation by inhibiting miR-338-3p and thereby upregulating many osteogenic-related genes [45]. Similarly, circ-RBM23 acted as a suppressor of miR-338-3p, enhancing RUNX2 and regulating the switch between osteogenesis and adipogenesis in MSCs. The authors reported that circ-RBM23 was elevated in osteogenesis, whereas it was depressed during adipogenesis in MSCs, providing a crucial goal for diagnosing and alleviating osteoporosis [46]. Furthermore, circ_0011269, circ-VANGL1, and circ_0076690 were notably decreased in the clinical samples of osteoporotic patients. In hMSCs, circ_0011269, circ-VANGL1, and circ_0076690 upregulated RUNX2 expression via sponging miR-122, miR-217, and miR-152, respectively. These findings pointed out that the overexpression of these circRNAs promoted osteogenic differentiation and could be implicated in the pathogenesis of osteoporosis [47,48,49].
Previous reports have revealed the functions of various circRNAs in influencing SMAD5 expression, making them potential targets for therapeutic interventions related to skeletal disorders. Circ_0001825 is newly recognized to be significantly downregulated in osteoporotic patients. The suppression of circ_0001825 reduced hMSC viability and osteogenic differentiation. Circ_0001825 promoted osteogenesis via sponging miR-1270, resulting in SMAD5 overexpression, which signifies the potential contribution of circ_0001825 to osteoporosis [50]. Another circRNA that has been detected to regulate SMAD5 in osteoporosis is CircGLIS2. The expression of circGLIS2 was obviously decreased in osteoporotic patients, while increased in hBMSCs upon osteogenic differentiation. Regression of circGLIS2 notably inhibited osteogenesis-related genes, such as osteopontin (OPN), OCN, and ALP activity. The study reported that circGLIS2 sponged miR-214-3p and upregulated Smad5, which stimulated osteogenic differentiation of the hBMSCs [51]. Circ_0062582 was also observed to lessen the development of osteoporosis by affecting SMAD5 expression. It was reported that circ_0062582 was remarkably decreased in osteoporosis. Meanwhile, circ_0062582 was increased in osteoblast medium-induced hBMSCs, where it sequestered miR-197-3p and resulted in an elevation of SMAD5, thereby reflecting hBMSC proliferation and osteogenic differentiation [52].
In vitro studies using MSCs revealed that circRNAs play a regulatory role on several genes related to the BMP pathway. Both circRNA_0048211 and circ_0016624 were suppressed in PMOP patients compared to healthy controls. During hMSC osteogenic differentiation, the upregulated circRNA_0048211 and circ_0016624 increased BMP2 regulation through targeting miRNA-93-5p and miR-98, respectively, so attenuated development of PMOP [53, 54].
Regarding the Notch pathway, circRNA_0001795 and circ-ITCH promoted osteogenic differentiation of hBMSCs by targeting YAP1 through sponging their respective miRNAs, miR-339-5p and miR-214 [55, 56]. Notably, the expression levels of both circRNAs were notably reduced in osteoporosis, and their depletion correlated with a decrease in the expression of osteogenic-related genes. Additionally, in vivo experiments demonstrated that the upregulation of circ-ITCH enhanced osteogenesis in ovariectomized (OVX) mice.
It has been found that hsa_circ_0114581 is negatively correlated with osteoporosis pathogenesis. The upregulated hsa_circ_0114581 enhanced the expression of heterogeneous nuclear ribonucleoprotein A3 (HNRNPA3) to encourage osteogenic differentiation by suppressing hsa-miR-155-5p. This study also proved that HNRNPA3 was directly associated with osteogenic-related proteins in BMSCs and femur samples of either human bone tissue or OVX mice, suggesting its relationship with osteogenesis and bone formation [57]. Moreover, low levels of circ_0019693 were recognized in osteoporotic patients. Its expression was further enhanced during the stages of osteogenic differentiation in BMSC. Intriguingly, it was indicated that circ_0019693 could promote osteogenesis through inhibiting miR-942-5p and increasing purkinje cell protein 4 (PCP4) [58]. When PCP4 is bound to CaM-dependent protein kinase, the rate of Ca+2 dissociation from calmodulin (CaM) increases, promoting calcium deposition during mineral nodule formation, which is a valid predictor of bone-forming ability [59]. Hsa_circ_0006215 is another circRNA coupled between osteogenesis and angiogenesis and involved in the pathogenesis of senile osteoporosis. It was significantly downregulated in the BMSCs of osteoporotic patients, while its overexpression promoted the osteogenic differentiation of BMSCs. In vivo, its overexpression induced the repair of the single cortical bone defect model, suggesting that it could promote bone defect repair. Furthermore, it has been demonstrated that hsa_circ_0006215 influences RUNX2 and VEGF expression in BMSCs through sponging miRNA-942-5p [60]. In GIOP patients, the hsa_circ_0006393 level was downregulated compared to control ones. Furthermore, overexpression of hsa_circ_0006393 boosted the expression of osteogenic genes in bone tissue samples obtained from male GIOP patients as well as a GIOP mouse model. It was suggested to have osteogenic influence through inhibiting miR-145-5p and increasing FOXO1 expression [61].
Numerous studies have been conducted on human adipose stem cells (ASCs). It was found that CircFOXP1 was significantly downregulated in the bone tissues of osteoporotic patients. Indeed, the pro-osteogenic activity of circFOXP1 was evaluated in vivo and in vitro, where circFOXP1 was found to enhance hASC osteogenesis by sponging miR-33a-5p and targeting FOXP1 expression [62]. Thus, circFOXP1 could be regarded as a candidate target for hASC-based therapy of osteoporosis.
Accumulating data indicates that circRNAs have the ability to interact with RBPs and alter their functions [63]. CircStag1 was remarkably suppressed in the BMSCs of osteoporotic rats and bone tissue samples isolated from osteoporotic patients. Briefly, circStag1 interacted with human antigen R (HuR) and promoted its translocation into the cytoplasm. Sequentially, cytoplasmic HuR led to the stimulation of the Wnt cascade by stabilizing low-density lipoprotein receptor-related protein 5/6 (Lrp5/6), co-receptors for Wnt ligands, and enhancing β-catenin expression. It was also observed that the induction of circStag1 fostered osteogenesis in OVX mice [64]. According to Yao et al., circ-Plod2 is an exon-type circRNA found in the cytoplasm, where it destabilized the Mpo-dependent osteogenic differentiation of BMSCs while having no effect on adipogenic differentiation or chondrogenic differentiation of these cells [65]. The study also found that the expression of circ-Plod2 was remarkably downregulated in OVX rats BMSCs, and its overexpression effectively lessened osteoporosis among them. Circ-Plod2 mediated its effect by interacting with IGF2BP2 to form an RNA–protein complex, which in turn inhibited the expression of Mpo mRNA in BMSCs. Another circRNA that plays a role in osteoporosis by interacting with RBP is circPVT1. CircPVT1 stimulates osteogenesis by inhibiting miR-30d-5p to increase integrin beta-3 (ITGB3) expression [66]. ITGB3, a member of the integrin family, functions as a downstream gene of Homeobox D3, which triggers the Wnt/β-catenin signaling pathway through the involvement of β3 integrin.
CircRNAs that inhibit osteogenesisOther circRNAs exert a pivotal regulatory role in impeding osteogenesis through modulation of essential pathways and gene expression networks, thereby influencing the overall process of bone formation. A recent study has highlighted the mechanistic insight of hsa_circ_0006859 in vitro. It was recognized that the expression of hsa_circ_0006859 was augmented in OVX mouse-derived BMSCs but remained modest expressed during osteogenic differentiation. Hsa_circ_0006859 overexpression hindered osteogenesis of BMSCs in the human fetal osteoblast cell line by targeting both miR-642b-5p/ephrin A2 (EFNA2) and miR-483-3p/dedicator of cytokinesis 3 (DOCK3) axes, resulting in the inactivation of the Wnt-signaling pathway [67]. Additionally, it has been reported that hsa_circ_0008842, identified as circZNF367, was substantially upregulated in the bone tissue of osteoporotic patients and dramatically downregulated in hBMSCs during osteogenic differentiation. The overexpression of circZNF367 suppressed migration and osteogenic differentiation of hBMSCs, both in vitro and in vivo, while its knockdown exhibited opposite effects. CircZNF367 could reduce osteogenic differentiation of hBMSCs through interaction with HuR, which reduced LRP5 mRNA stability [68].
Circ_0006873 was upregulated in the sera of osteoporotic patients and decreased during osteoblastic differentiation. Circ_0006873 could suppress osteoblastic differentiation and favor osteoporosis by sponging miR-142-5p, which in turn enhances PTEN expression and regulates the Akt signaling pathway [69]. Another study has also reported that circ_0006873 was significantly elevated in both serum samples and bone tissue samples of osteoporotic patients. Its overexpression was associated with a significant reduction in osteogenic differentiation of hMSCs through the sequestration of miR-20a and targeting SMURF2 [70].
Lipopolysaccharide (LPS) is a potent inducer of bone loss, resulting in inflammation that exacerbates osteoporosis by disrupting regular bone homeostasis. CircAtp9b is LPS-inducible, and its knockdown alleviates the inflammation triggered by LPS [71]. Interestingly, circAtp9b was significantly upregulated in plasma samples and osteoblasts derived from osteoporotic patients. LPS-treated osteoblasts increased circAtp9b expression in a dose-dependent manner, confirming that the circAtp9b overexpression is likely triggered by LPS. Also, it was shown that circAtp9b upregulation increased LPS-induced osteoblast apoptosis in osteoporosis by sponging premature miR-17-92a and suppressing its maturation, which in turn accelerated osteoporosis progression [72].
There is little known about the role of circRNAs in melatonin (MEL)-induced BMSC osteogenic differentiation and osteoporosis progression. MEL has been detected as a booster of osteoblast proliferation and differentiation, fostering bone formation and reducing bone destruction in osteoporotic mice [73]. In this regard, Wang et al. found that melatonin improved osteogenic differentiation and repressed osteoporosis development by hampering circ_0003865 expression [74]. This circRNA acts as a sponge for miR-3653-3p, consequently boosting the expression of growth arrest-specific gene 1 (GAS1) while suppressing the expression of bone-forming genes. Similarly, the expression of circ_0005753 was significantly reduced during the osteogenic differentiation of BMSCs triggered by MEL. Molecularly, circ_0005753 maintained the stability of TXNIP mRNA through the recruitment of PTBP1. This study pointed out that MEL enhanced BMSC osteogenic differentiation through the regulation of the circ_0005753/PTBP1/TXNIP axis, which could shed light on a new treatment pathway to prevent osteoporosis [75].
Zhi et al. determined that exosomal hsa_circ_0006859 was obviously upregulated in the serum of PMOP, efficiently distinguishing osteoporotic and osteopenic patients from healthy controls [76]. As well, it discriminated between osteoporotic and osteopenic patients with high specificity and sensitivity. Besides, ectopic expression of circ_0006859 provoked adipogenic differentiation and impeded osteoblastic differentiation in BMSCs via sponging miR-431-5p to induce ROCK1 expression. Thus, hsa_circ_0006859 may be a potential biomarker for the diagnosis and prognosis of osteoporosis and could modulate the harmony between adipogenesis and osteogenesis in BMSCs. According to whole transcriptome sequencing, Zhang et al. determined the expression profile of circRNAs in the peripheral blood of male osteoporotic patients versus healthy controls, and found that a total of 398 circRNAs were differentially expressed [77]. Hsa_circ_0042409 was among the top 10 upregulated circRNAs and was recognized to be involved in the development of osteoporosis through regulating the expression level of kinesin light chain 1 (KLC1) via sponging hsa-miR-195-5p. Additionally, another study observed a significant upregulation of circ_0134944 in both PMOP patients and OVX mice. Its enhanced expression suppressed osteogenesis in BMSCs through targeting miR-127-5p, causing an increase in pancreatic and duodenal homeobox 1 (PDX1) and sphingosine kinase 1 (SPHK1). PDX1 is a well-established transcription factor that regulates the activity of multiple genes by binding to their promoter regions, such as SPHK1, which has an impact on the regulation of osteogenic differentiation [78]. Furthermore, Hsa_circ_0002060 knockdown in the human fetal osteoblast cell line (hFOB1.19) reversed the effect of DEX, which decreased matrix metalloproteinases (MMP) and increased ROS. Hsa_circ_0002060 modulated the survival of hFOB 1.19 cells by targeting miR-198-5p, resulting in an elevation of Bax expression that in turn triggered the apoptosis of osteoblasts. Additionally, hsa_circ_0002060 knockdown alleviated the progression of osteoporosis in OVX mice through the Jun N-terminal kinase (JNK) signaling pathway [79].
Collectively, manipulating these interrelated molecules could be beneficial in the management of osteoporosis. Thus, we reviewed the expression and molecular mechanisms by which circRNAs mediate osteoblast differentiation (Table 2 and Fig. 3).
Table 2 The expression of circRNAs-related osteogenesis in osteoporosisFig. 3
Role and mechanistic insights of circRNAs in osteogenesis. All shapes shaded in green denote biomarkers increased during osteogenesis. All shapes shaded in red denote biomarkers decreased during osteogenesis
Osteoclastogenic signaling pathwaysOsteoclastogenesis is the fusion of osteoclast precursors, which originate from hematopoietic cells to form multinucleated, active osteoclasts. Macrophage colony-stimulating factor (M-CSF), also known as CSF-1, and receptor activator for nuclear factor κB Ligand (RANKL) are two crucial cytokines that bind to their respective receptors, colony-stimulating factor-1 receptor (c-Fms) and RANK, to stimulate osteoclastogenesis through the regulation of delicate signaling pathways [80]. M-CSF stimulates the production and survival of osteoclast precursor cells by activating extracellular signal-regulated kinase (ERK) and serine/threonine kinase 1 (Akt). Meanwhile, RANKL facilitates the development of osteoclast precursors into osteoclasts by attracting TRAF6 to the RANK receptor, which then activates various downstream targets, including NF-κB, JNK, ERK, p38, and PI3K/Akt. This ultimately leads to the activation of the key regulator of osteoclast differentiation, nuclear factor of activated T cells 1 (NFATc1). This latter controls several osteoclast-specific genes, including TRAP, cathepsin K, and osteoclast-associated receptor (OSCAR) via interaction with other osteoclastic transcription factors, such as microphthalmia-associated transcription factor (MITF) and c-Fos [81]. Additionally, the RANK/RANKL/OPG pathway is crucial for the metabolism of bone tissue, as RANKL stimulates the production and differentiation of osteoclasts, while osteoprotegerin (OPG) inhibits its action through binding to it [82]. Here, we summarized the relevant research investigation of lncRNAs (Table 3) and circRNAs (Table 4) in order to offer novel insights for future osteoporosis studies and treatments, with a specific focus on osteoclasts, as depicted in Fig. 4.
Table 3 The expression of lncRNAs-related osteoclastogenesis in osteoporosisTable 4 The expression of circRNAs-related osteoclastogenesis in osteoporosisFig. 4
留言 (0)