
記住我
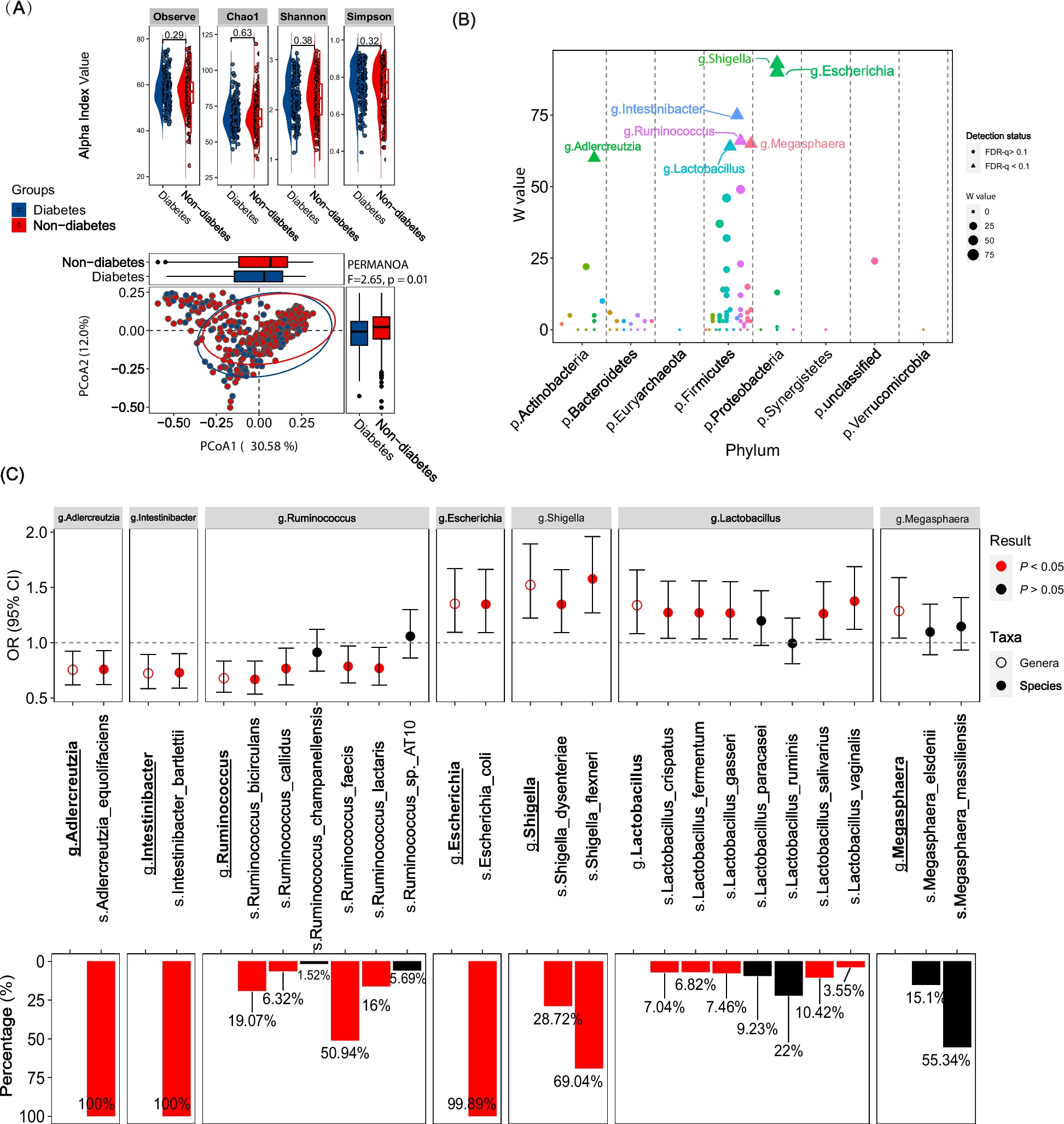


The clinical features of all 163 enrolled patients were examined and a side-by-side comparison among different cohorts was performed. Majority of patients (97.1%, 136 out of 140 patients whose treatment information is accessible) received first-line asparaginase-based chemotherapy. The NODE and ICGC cohorts are mostly comprised of primary patients, whereas the NKT_project and EGA cohorts contain more relapsed/refractory patients (Additional file 2: Table S2). Their prognoses vary among different cohorts (Additional file 3: Fig. S1). To evaluate the impacts of different clinical features on the overall survival, we performed both univariate and multivariate Cox regression analyses. While the analysis on different disease stages, primary vs. relapsed/refractory status, prognostic scores, and patient cohorts all appear as significant in the univariate analysis, only the relapsed/refractory status is significantly associated with poor prognosis in the multivariate analysis (Additional file 2: Table S3). Patients with relapsed/refractory NKTCL generally show more advanced Ann-Arbor stage and higher IPI and PINK scores compared with their primary counterparts (Additional file 2: Table S4). The overall survival (OS) of relapsed/refractory patients is significantly worse in comparison (5-year OS rate 57.9% versus 27.4%, P = 0.003, Additional file 3: Fig. S1).
For primary patients (n = 127), a total of 6826 nonsilent somatic mutations were identified, including 5904 missense mutations, 373 nonsense mutations, 340 out-frame INDELs, and 177 in-frame INDELs. For relapsed/refractory patients (n = 36), we identified a total of 3512 nonsilent mutations, comprising 2503 missense mutations, 254 nonsense mutations, 392 out-of-frame INDELs, and 366 in-frame INDELs. The per-sample mutation count in relapsed/refractory patients is significantly higher than that in the primary patients (per-sample median: 68 vs. 48, Brunner–Munzel test, P = 0.003, Fig. 1A, B). Likewise, a higher tumor mutation burden (TMB) level was also found in relapsed/refractory patients (Brunner–Munzel test, P = 0.003, Fig. 1C, D). To account for the potential confounding factor introduced by sequencing method difference (i.e., WES vs. WGS), we conducted separate analyses for patients with WES and WGS data. For patients with WES data, relapse/refractory patients show a median count of 55 nonsilent somatic mutations, compared to 40 in the primary group (Additional file 3: Fig. S2). Additionally, the relapse/refractory patients exhibit significantly higher levels of TMB than the primary group (per-sample median: 1.1 vs. 0.8 per megabase, Brunner–Munzel test, P = 0.022, Additional file 3: Fig. S2). For patients with WGS data, we also observed a higher median count of nonsilent somatic mutations in the relapse/refractory group (per-sample median: 103 vs. 60, Additional file 3: Fig. S2) compared with the primary group. Significant disparity also exists at the TMB level (per-sample median: 2.1 vs. 1.2 per megabase, Brunner–Munzel test, P = 0.017, Additional file 3: Fig. S2). Taken together, these results suggest a higher nucleotide-level mutational load in relapsed/refractory NKTCL patients. In addition, we also noticed that the relapsed/refractory patients exhibit significantly higher MSI scores compared to their primary counterparts (per-sample median: 0.755 vs. 0.120, Brunner–Munzel test, P = 0.001, Fig. 1E). Such differences remained as significant when considering patients with WES and WGS data separately (Additional file 3: Fig. S2). This observation further highlights the higher levels of genome instability in the relapsed/refractory NKTCL patients compared with their primary counterparts.
Fig. 1
Characterization of somatic mutations and tumor mutation burden in NKTCL. A The per-sample numbers of different categories of non-synonymous somatic mutations in primary (left) and relapsed/refractory (right) NKTCL. B Per-sample distribution of different categories of non-synonymous mutations in patients with primary (left) and relapsed/refractory (right) NKTCL. The dashed line denotes the median value. C The tumor mutation burden (TMB) profiles of primary (left) and relapsed/refractory (right) patients. The dashed line denotes the median value. D The comparison of TMB levels between the primary and the relapsed/refractory group. Brunner–Munzel test was applied for assessing the statistical significance. E The comparison of MSI score between the primary and the relapsed/refractory group. Brunner–Munzel test was applied for assessing the statistical significance
Enrichment of mutational signatures associated with DNA damage repair defection in relapsed/refractory NKTCL patientsWe next examined the spectrum and context of SNVs. We saw a strong signal of C- > T transition that was also observed in other solid tumors [34], suggesting possible involvement of epigenetic alterations like DNA methylation in the pathogenesis of NKTCL. Three prominent single base substitution (SBS) signatures popped up among all samples: SBS1, SBS5, and SBS6, all of which directly contribute to C- > T mutations (Fig. 2A). Etiologically, SBS1 is explained by spontaneous or enzymatic deamination of 5-methylcytosine (m5C) to thymine [35]. SBS5 often correlates with SBS1, with potential link to tobacco smoking [36]. SBS6 is frequently observed in microsatellite unstable tumors with association to DNA repair defection [36]. We also analyzed mutational signatures of INDELs and identified the ID8 signature as the most dominant signature (Fig. 2B). ID8 is related to the repair of double strand breaks (DSBs) by non-homologous DNA end-joining (NHEJ) and bears similarity to radiation induced mutations [36, 37]. Notably, both SBS6 and ID8 are especially enriched in patients with relapsed/refractory NKTCL (Fig. 2C). Given that these two signatures are both associated with defective DNA damage repair, this observation hints that genes responsible for DNA damage repair might be a common mutational targets in relapsed/refractory NKTCL patients, which also explains the higher mutational load observed in these patients.
Fig. 2
Mutational signatures operating in NKTCL. A Major SBS signatures (SBS1, 5, 6) identified from the 163 NKTCL patients. B Major INDEL signature (ID8) identified from the 163 NKTCL patients. C Proportions of mutations bearing the SBS6 and ID8 signatures in the primary and relapsed/refractory groups respectively. Brunner–Munzel test was applied for assessing the statistical significance
Shared and distinct coding mutations between primary and relapsed/refractory NKTCLRegarding common mutated gene targets in NKTCL, we compiled a short list of recurrent mutated genes by filtering out those genes mutated in < 5 patients or frequently appeared in non-disease-specific WES datasets [38] (Fig. 3A and Additional file 3: Fig. S3). Consistent with previous reports [39], STAT3 comes up as the most frequent mutated gene (28%) with most mutations occurred in its SH2 domain (47 of 53 mutations) (Additional file 3: Fig. S3B). Other genes of the JAK-STAT signaling pathway (e.g., CBL and JAK3) are also frequently mutated. We detected TP53 mutations in 16% of patients, with the mutated sites predominantly located in its P53 DNA-binding domain (Additional file 3: Fig. S3C). Of the helicase gene family, we recaptured the RNA helicase gene DDX3X (18%) as a recurrent mutation target, with various missense mutations affecting its ATP-binding helicase domain (Additional file 3: Fig. S3D). Meanwhile, mutations in another helicase gene HELZ2 are also detected in 13% of patients. Other identified recurrent mutated genes include the following: tumor suppressors (CSMD1, 18%; ZFHX3, 17%; FAT1, 16%; FAT4, 12%; KMT2C, 11%; MYO18B, 11%; NOTCH1, 10%; SLIT2, 9%; NOTCH2, 7%; SLX4, 7%), genome instability modulators (DST, 23%; ALMS1, 16%; PCNT, 14%; PRDM9, 11%; PRKDC, 10%; DOCK4, 10%; TUBGCP6, 7%; NF1, 7%), and epigenetic modifiers (ARID1A, 14%; TET2, 11%; KDM6B, 11%; HDAC9, 9%; KDM2B, 8%; ARID1B, 7%).
Fig. 3
Mutational landscape of primary and relapsed/refractory NKTCL. A The top panel shows the frequency of non-synonymous mutations in each sample. The main panel shows the occurrence of major somatic mutations across different samples, with different colors correspond to different mutation categories. Genes involved in these mutations are grouped based on their biological functions indicated on the left. The overall frequency of these somatic mutations across all examined samples are shown on the right. The lower panel indicates the sampling status (primary vs. relapsed/refractory), sequencing methods (WGS vs. WES), and Ann Arbor stage (I/II vs. III/IV) of each sample. B The co-occurrence and mutual exclusivity patterns among the frequent mutations. The font colors of gene names represent the biological function depicted in (A). C Kaplan–Meier curves of overall survival (OS) by the mutation status of TP53, PRDM9, and STAT3. D Kaplan–Meier curves of OS by the mutation status of TP53-STAT3 and KMT2C-NF1. E The mutational landscape of somatic mutations with significant higher frequency in relapsed/refractory patients than in primary patients (Fisher’s exact test, P < 0.05). The mutational frequency of these genes in primary patients is shown on the left, while their corresponding mutational frequencies in relapsed/refractory patients are shown on the right. The mutated genes are ranked by the mutational frequency in the latter group
We further investigated the co-occurrence and mutual exclusivity among these common mutations in NKTCL. We found 44 gene pairs showing significant co-occurrence mutations (Fisher’s exact test, P < 0.05), with NOTCH1-KDM6B (P = 0.001), CBL-KDM2B (P = 0.001), and DST-HELZ2 (P = 0.001) being on the top of the list (Fig. 3B). A previous study highlighted KDM6B’s crucial role in supporting NOTCH1-driven T cell acute lymphoblastic leukemia [40]. The synergistic effect of NOTCH1 and KDM6B mutations therefore may accelerate lymphoma progression by activating oncogenic pathways via chromatin remodeling. Other interactions also likely exacerbate oncogenic stress by fostering lymphoma cell proliferation. Future studies are needed to evaluate the therapeutic potential of targeting these interactions. To explore the prognostic value of these recurrent mutations and their co-occurring combinations, we applied the Kaplan–Meier analysis and discovered that mutations in TP53, PRDM9, and STAT3 are significantly associated with worse OS (Fig. 3C). While such association has been previously reported for TP53 and STAT3 [39, 41], our study provides the first evidence for the prognosis value of PRDM9 aberration. We found patients with STAT3-TP53 co-mutations exhibited a worse prognosis than patients with a single or no mutation of STAT3 and TP53 (P = 0.004) (Fig. 3D). Similarly, patients carrying KMT2C-NF1 co-mutations are significantly associated with poor prognosis compared to those carrying either a single mutation or no mutation in these two genes (P = 0.03) (Fig. 3D).
We found 68 genes that are significantly more likely to be mutated in relapsed/refractory patients (Fisher’s exact test), which include genome instability modulators (CDC27, P = 0.002; PRPF4B, P = 0.006; PRDM9, P = 0.009; NF1, P = 0.015) and epigenetic modifiers (ARID1B, P = 0.015; HDAC9, P = 0.009; NCOR1, P = 0.040) as well as tumor suppressors (CUX1, P = 0.001; ZFHX3, P = 0.007; PTPRK, P = 0.008; CBL, P = 0.013; TP53BP1, P = 0.043) (Fig. 3E). Given that relapsed/refractory patients are generally associated with more advanced Ann Arbor stages (Fisher’s exact test, P < 0.001), it is likely that these differentially mutated genes play important roles in promoting the progression of NKTCL. We also examined the co-occurrence and mutual exclusivity among these 68 genes, which reveals a series of co-occurrence mutations (Fisher’s exact test, P < 0.05, Additional file 3: Fig. S3E) such as ZFHX3-FNDC1 (P < 0.001), HDAC9-FGFR1 (P = 0.002), and ACACA-NF1 (P = 0.02). The prevalence of these co-occurring mutations underscores the role of synergistic mutations in driving the progression and evolution of NKTCL.
In addition, the infection status of Epstein–Barr virus (EBV) has also been reported to be significantly associated with the prognosis of NKTCL [42]. For the patients with accessible EBV-infection data (n = 115), we evaluated the correlation between patients’ peripheral blood EBV-infection status and their NKTCL mutational profiles. For those most-frequently mutated genes in NTKCL such as STAT3, DST, CSMD1, DDX3X, no significant difference can be detected regarding their mutational frequency between the EBV-positive and EBV-negative groups. However, specific gene mutations are notably enriched in the EBV-positive group (Fisher’s exact test), including UBR4 (P = 0.010), C2orf16 (P = 0.019), CACNA1E (P = 0.036), UTP20 (P = 0.014), and COL6A5 (P = 0.030). Conversely, certain genes such as HERC2 (P = 0.047) and RRP1 (P = 0.018) are significantly enriched in the EBV negative group (Additional file 3: Fig. S3F). UBR4 has been reported to interact with viral proteins that are important for viral life cycle [43]. In EBV-positive NKTCL, frequent mutations in UBR4 could influence the viral manipulation of host cellular mechanisms, potentially leading to oncogenic processes such as immune evasion and unregulated cellular proliferation. The positive association between UBR4 mutations and EBV infection underscores the role of UBR4 in the pathogenesis of EBV-associated malignancies and highlights its potential value for therapeutic intervention.
Higher copy number alteration and aneuploidy occurrences in relapsed/refractory patientsWe profiled the somatic CNVs landscape based on both WGS and WES data. The number of CNVs vary substantially across different samples, with an average of 138.2 CNVs identified per sample (Fig. 4A). We classified these CNVs into four categories: single CN loss (CN = 1 copy), double CN loss (CN = 0), low-level CN gain (CN = 3), and high-level CN gain (CN > 3 copies). Nearly 90% of CNVs are single CN loss (42.5%) and low-level CN gain (44.8%), which is different from previous observations in other solid tumors where ~ 52.4% cases were detected with amplicons of CN ≥ 5 [44]. We found a significantly higher CNV burden in relapsed/refractory patients (per-sample median: 58.5 vs. 33, Brunner–Munzel test, P = 0.008, Fig. 4B, C), reflecting their elevated genome instability. Consistent results were obtained when analyzing the WGS and WES datasets separately (Additional file 3: Fig. S4). Aneuploidy is an extreme type of CNV that spans over entire chromosome arms with substantial impact on genome instability. We observed a significantly higher aneuploidy level for relapsed/refractory patients (median score: 9) than their primary counterparts (median score: 5) (Brunner–Munzel test, P < 0.001, Fig. 4D).
Fig. 4
The landscape of copy number variation in NKTCL. A Distribution of different categories of CNVs across the 163 NKTCL patients. The CNV categories employed here include single CN loss (CN = 1 copy), double CN loss (CN = 0), low-level CN gain (CN = 3), and high-level CN gain (CN > 3 copies). B Heatmap of CNVs in patients with primary and relapsed/refractory NKTCL. C The per-sample CNV number comparison between the primary and the relapsed/refractory groups. Brunner–Munzel test was applied for assessing the statistical significance. D The per-sample aneuploidy score comparison between the primary and the relapsed/refractory groups. Brunner–Munzel test was applied for assessing the statistical significance. E KEGG pathway enrichment analysis of gene-level CNVs for patients with NKTCL. F Gene-level CNV landscape across the 163 NKTCL patients
At the gene-level, we identified several genes with recurrent CN changes. These genes are associated with biological function such as genome integrity (HMGB3, PMS2, GPX4, DNMT1, CDKN2D, and TP53), JAK-STAT signaling (CRLF2, TYK2, EPOR, STAT3), tumor suppressors (RBMX, DUSP9, STK11, AMH, TCF3, PIN1, NOTCH3), and epigenetic modifiers (MECP2, CARM1, DOT1L). At the pathway-level, these genes are significantly enriched in pathways involved in EBV infection, JAK-STAT signaling, and DNA replication and mismatch repair (Fig. 4E). When comparing the CNV profiles between primary and relapsed/refractory patients, we found genes such as CRLF2, PMS2, and STK11 show significantly higher propensity for copy number alterations in relapsed/refractory patients (Fig. 4F). Among these genes, the frequency of CRLF2 CN gain in relapsed/refractory patients is significantly higher than that in the primary group (21/36 vs. 8/127, P < 0.001). CRLF2 overexpression has been widely confirmed to be associated with adverse outcomes in B cell acute lymphoblastic leukemia (B-ALL) [45]. It has been shown that high-dose of thymic stromal lymphopoietin (TSLP) can induce apoptosis, impeding the proliferation and migration of CRLF2 B-ALL tumor cells [46]. Furthermore, the synergistic combination of the JAK1/2 kinase inhibitor ruxolitinib with conventional treatment has demonstrated enhanced efficacy in CRLF2-altered B-ALL [47]. These findings offer valuable insights into potential treatment options for patients with relapsed/refractory NKTCL.
By leveraging the exact copy number, segment size, and heterozygosity status of these CNVs [48], we summarized the CN signatures for each NKTCL sample (Additional file 3: Fig. S5A). We found CN1, CN2, CN5, and CN9 are among the most prominent signatures. Notably, CN2 (tetraploid genome, i.e., whole genome doubling), CN5 (chromothripsis), and CN9 (chromosomal instability in diploid genome) are all associated with genome instability [49]. In particular, CN9 is significantly more predominant in patients with relapsed/refractory NKTCL (Brunner–Munzel test, P = 0.041; Additional file 3: Fig. S5B). Patients with the CN9 signature show significantly inferior OS (Additional file 3: Fig. S5C).
Landscape of canonical and complex structural rearrangements in NKTCL patientsFor NKTCL patients whose WGS data is available, we next extend our analysis to canonical SVs such as insertions, deletions, duplications, inversions, and translocations. Altogether, we identified a total of 6131 events across all 71 NKTCL patients (mean: 86.3 per patient) with 29% of them being segmental deletions (Fig. 5A). Consistent with our observations based on SNV, INDELs, and CNVs, we found the overall SV burden of relapsed/refractory patients is significantly higher than that of primary patients (Brunner–Munzel test, P < 0.001, Fig. 5B). Also, patients with TP53 mutations show more SVs than those without TP53 mutations (Brunner–Munzel test, P = 0.002), highlighting the impact of TP53 inactivation on SV accumulation (Fig. 5C).
Fig. 5
The landscape of canonical and complex structure variation in NKTCL. A Distribution of canonical structural variants (SVs) across the 71 NKTCL patients with WGS data. The canonical SV types employed here include insertion, deletion, duplication, inversion, and translocation. B The comparison of per-sample canonical SV numbers between the primary and relapsed/refractory patient groups. Brunner–Munzel test was applied for assessing the statistical significance. C The comparison of per-sample canonical SV numbers between the wildtype and mutated TP53 patient groups. Brunner–Munzel test was applied for assessing the statistical significance. D An exemplary chromothripsis event detected in one NKTCL patient. The chromosome ideograms of the affected regions (on chromosomes 1 and 2) are shown at the top. Different subtypes of SVs are represented by connected lines illustrated in the middle: black, head-to-head inversion (+ / +); green, tail-to-tail inversion (− / −); orange, deletion-like (+ / −); blue, duplication-like (− / +). The copy number profile of the corresponding regions is depicted at the bottom, where the horizontal and vertical axes represent the chromosome coordinates and copy number respectively, while each CN segment is represented by a black bar. Detail information for the depicted region is presented as a table at the bottom. E Kaplan–Meier curves of progression-free survival (PFS) by the attribute of chromothripsis in the NKTCL patients with WGS data. F Kaplan–Meier curves of overall survival (OS) by the attribute of chromothripsis in the NKTCL patients with WGS data. G Detailed characterization of 12 samples identified with focal amplifications, for which focal amplicons were further classified into linear, complex, cyclic (eccDNA), and breakage fusion bridge (BFB). H The genome browser track derived from one patient (denoted as NKT_44) depicted by AmpliconArchitect. The connected lines indicate potential structure combinations in eccDNA amplicons. The horizontal axis represents chromosome coordinates. The left vertical axis represents the depth of coverage, while the right vertical axis denotes the copy number
SVs can promote tumorigenesis and progression by forming fused oncogenes or by disrupting tumor suppressor genes [50]. Indeed, we identified a set of cancer-related genes recurrently interrupted by SV breakpoints (Additional file 2: Table S5). For example, CD274 (PD-L1) encodes an immune inhibitory receptor ligand, and its expression is associated with tumor immune escape in many types of cancers [51]. We identified SVs in this gene in 17.2% of primary and 7.7% of relapsed/refractory patients respectively, with 38.4% of such SV cases being tandem duplications. LRP1B disruption has been shown as a potential contributor to the emergence of chemotherapy resistance in ovarian cancer [52]. Our findings reveal that LRP1B is also frequently interrupted by SV breakpoints in NKTCL, notably to a much larger extent in relapsed/refractory patients (6.9% in primary versus 30.8% in relapsed/refractory; Fisher’s exact test, P = 0.034). Therefore, the higher SV burden of LRP1B in NKTCL patients, especially those relapsed/refractory ones, likely serves to reduce chemotherapy sensitivity and therefore to lead to treatment failures. In addition, we found RBFOX1 translocations also occur much more frequently in relapsed/refractory NKTCL patients (1.7% in the primary vs. 30.8% in relapsed/refractory; Fisher’s exact test, P = 0.003). RBFOX1 promotes mRNA stability and it is down-regulated across multiple cancers types [53]. The frequent translocation observed in this gene in NKTCL is consistent with such pan-cancer pattern. This highlights the importance and potential therapeutic value for restoring post-transcriptional homeostasis in anticancer treatment.
We further extended our analysis to more complex SV types such as chromothripsis and eccDNA. We identified chromothripsis in 15 out of 71 patients (21.1%), with a significantly higher identification rate in the relapsed/refractory group (primary: 8/58 vs. relapsed/refractory: 7/13; Fisher’s exact test, P = 0.004). Chromosomes 1, 2, and 7 are frequently impacted by chromothripsis (Fig. 5D). TP53 mutation has been reported to be associated with chromothripsis in solid tumors [54]. Concordantly, we found 50% (3/6) of TP53-mutated NKTCL patients have chromothripsis, whereas only 18.5% (12/65) of those NKTCL patients with wild-type TP53 have chromothripsis, underscoring the risk of TP53 mutation in triggering nuclear catastrophe. Patients suffer from chromothripsis show significantly inferior PFS and OS, highlighting the role of chromothripsis in promoting NKTCL progression (Fig. 5E, F).
Recent studies reported that chromothripsis could serve as a precursor for focal amplifications, which further promote the rapid amplification of oncogenes [55]. We detected a total of 65 focal amplification events in 12 out of 71 NKTCL patients, with 7 of them also have chromothripsis. Such focal amplifications are much more common in relapsed/refractory patients (9/13) than in primary patients (3/58) (69.2% vs 5.2%, P < 0.001, Fisher’s exact test). The detailed count and subtype of focal amplifications are heterogeneous across different patients, with linear focal amplifications being the most dominant subtype (n = 55) (Fig. 5G). As a special subtype of focal amplifications, eccDNA has been reported to be an important oncogenic driver that also contributes to drug resistance [18, 56]. We found eccDNA in 3 out of 12 patients with focal amplifications. Remarkably, in one patient (denoted as NKT_44) with eccDNA, the JAK2, CD274 (PD-L1), and PDCD1LG2 (PD-L2) genes all got substantially amplified with eccDNA (Fig. 5H). JAK2 is a critical component of the JAK-STAT signaling pathway whose over-activation can lead to NKTCL, while the amplification of CD274 (PD-L1) and PDCD1LG2 (PD-L2) genes will foster immune escape. Therefore, for this particular case, both JAK-STAT over-activation and immune escape contribute to the pathogenesis of NKTCL.
Integration of genomic alterations of full spectrum to derive a novel molecular subtyping system for NKTCLGiven that we have systematically characterized genomic alteration landscapes of full spectrum (SNVs, INDELs, CNVs, and SVs) for primary and relapsed/refractory NKTCL patients, we further evaluated their prognostic values with NMF clustering [32]. A total of 151 patients were successfully classified into four clusters (C1–C4) (Fig. 6A). As for the remaining 12 patients, no characteristic driver genetic alterations were identified and therefore we classified them as the C0 cluster, although their genomes did show substantial alterations (mean SNVs = 26.4 per sample, mean CNVs = 63.6 per sample, mean SVs = 9.6 per sample) as those of other patients. All patients classified as C0 are primary patients. The C1 cluster is characterized by the mutations in DST, RELN, ARID1A, and CN loss of RBMX. The CD274 (PD-L1) SVs are also highly noticeable in this cluster. The C2 cluster is represented by the mutations involving the JAK-STAT pathway, including STAT3 and CN gain of CRLF2. The C3 cluster is associated with mutations in epigenetic modifiers such as TET2, KDM6B, and ARID1B as well as the mutations of tumor suppressors including FAT1 and KMT2C. Finally, the C4 cluster is characterized by the CN gain of STK11 and EPOR and mutations in genomic instability including PCNT, PRDM9, TUBGCP6, and NF1. Interestingly, patients from the C4 cluster predominantly have advanced-staged NKTCL with many of them being relapsed/refractory, whereas most patients from C1–C3 clusters have primary NKTCL (Fig. 6A). In accordance to this observation, the C4 cluster is also associated with the worst prognosis and higher PINK and IPI scores (Additional file 3: Fig. S6). Taken together, we found relapsed/refractory NKTCL patients are characterized by high levels of TMB, CNVs, and SVs, as well as higher propensity for chromothripsis and focal amplifications, reflecting their severe genome instability (Fig. 6B).
Fig. 6
Identification of clusters of NKTCL with coordinate genomic alterations. A Nonnegative matrix factorization clustering was carried out using somatic SNVs, INDELs, CNVs, and SVs in the 163 NKTCL patients (columns). Samples without candidate alterations were defined as cluster C0. Clusters C1–C4 with their associated representative genetic alterations are visualized. B Sankey-diagram of the clinical and mutational characteristics for NKTCL patients. The eight columns from left to right represent molecular subtyping clusters, Ann Arbor stage, sampling status, TMB, CNV, SV, chromothripsis, and focal amplifications respectively, with the total height represents the full 71 WGS samples. The curves with different colors show the correspondence relationship among different characteristics and molecular subtyping clusters. C Kaplan–Meier curve of PFS and D OS of all NKTCL patients from different molecular subtyping clusters. E Kaplan–Meier curve of PFS and F OS of primary NKTCL patients from different molecular subtyping clusters
With this molecular subtyping analysis, we mainly want to identify characteristic genomic alterations that helps to classify patients and potentially guide their treatment strategies, which is the ultimate goal of precision medicine. Nevertheless, it is also interesting to test if our molecular subtyping system captures the inherent factors that determines patients’ prognosis. First, we found patients from these five clusters (i.e., C0–C4) show clear differences in PFS (P < 0.001, Fig. 6C) and OS (P = 0.005, Fig. 6D). Moreover, by excluding relapsed/refractory patients and performing the survival analysis for the 127 primary patients only, we found our molecular subtyping systems is still capable of differentiating the prognostic performance of primary NKTCL patients in terms of both PFS (P = 0.004, Fig. 6E) and OS (P = 0.04, Fig. 6F). These results underscore the value of our molecular subtyping system for future clinical practice on NKTCL.
留言 (0)