
記住我

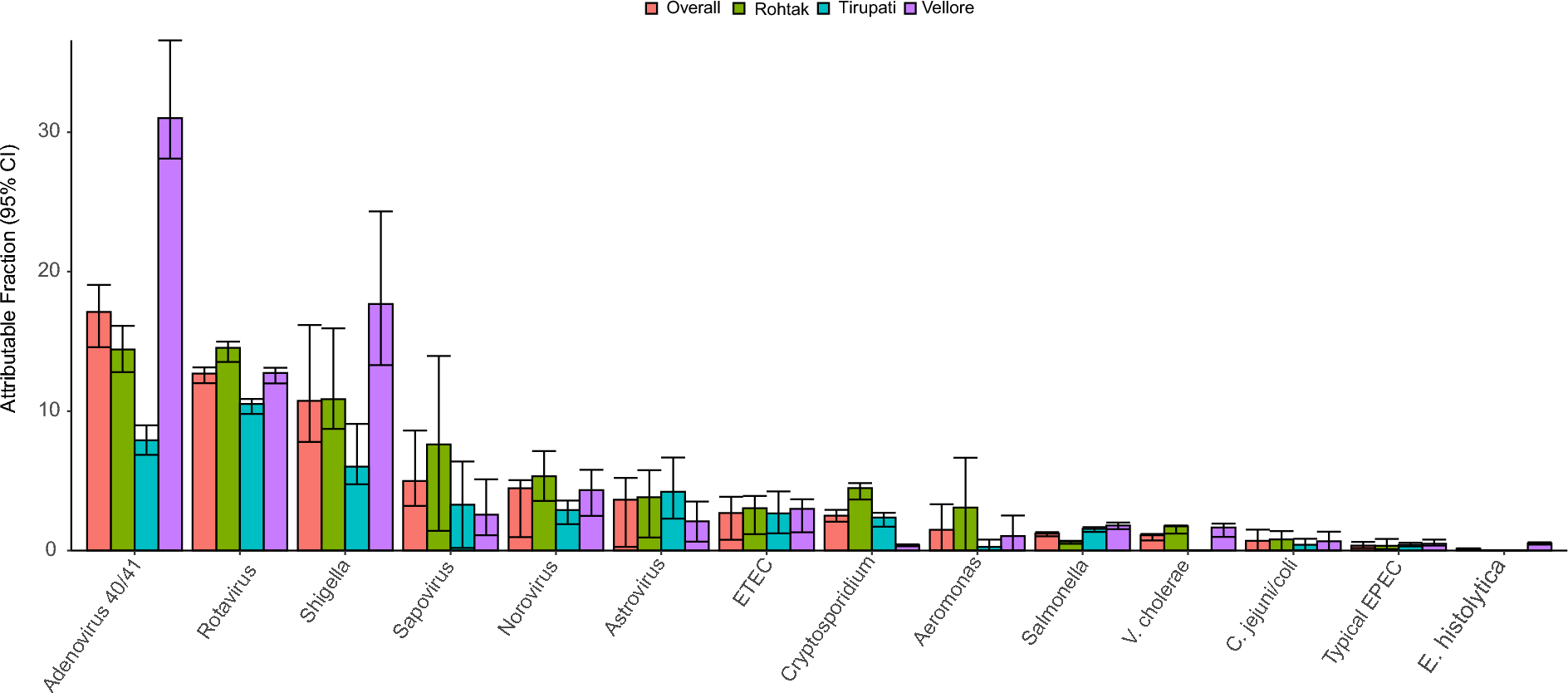

In this targeted LC–MS/MS metabolomics investigation, of initially 78 selected compounds of microbiome origin, we identified eight unique, statistically significant urinary metabolites at T1: 2-methylbutyrlglycine (2MBG), 3-hydroxypropionic acid (3HPA), 3-methylcrotonylglycine (3MCG), 4-hydroxyhippuric acid (4HHA), 5-hydroxyindoleacetic acid (5HIAA), 5-hydroxyhexanoic acid (5HHA), isobutyrylglycine (IG), and phenylacetylglutamine (PAG). Five of these eight significant metabolites (2MBG, 3MCG, 4HHA, 5HIAA, and PAG) showed a constant elevated level for TBM stage 1 (Fig. 4) across the entire treatment period and we consider this to be the background effect of the treatment. We consider these eight metabolites to be associated with the disease pathogenesis (TBM), as markers of the breakdown of M. tb cell wall caused by anti-TB medication, and/or dysbiosis commonly associated TBM. We discuss the biological significance of these eight metabolites in terms of: (1) associated metabolic pathways and corroboration from the literature; (2) their origin – linking these metabolites to either microbial or host origin, or a combination of both (a co-metabolite); (3) the insight(s) into the biology of the breakdown of M. tb cell wall within the host; (4) understanding the gut dysbiosis associated with TBM.
Metabolic map and link to previous TB marker studiesIt should be noted that seven of the eight significant metabolites identified in this study have previously been linked to pulmonary TB. IG, 3MCG and 2MBG are glycine conjugates of intermediates (isobutyryl-CoA, 3-methylcrotonyl-CoA and 2-methylbutyryl-CoA, respectively) of branched-chain amino acid catabolism (Fig. 5). Based upon a transcriptomics study by Jiang et al. [24], the expression of the enzyme branched-chain amino acid transferase-1 (BCAT1) is decreased, whereas BCAT2 increases, in TB cases. Both BCAT1 and BCAT2 are of human origin, with BCAT1 being localized in the cytoplasm and BCAT2 compartmentalized in the mitochondria. An increased expression of (mitochondrial) BCAT2 is in line with the enhanced energy metabolism associated with TB [8]. However, raised BCAT2 activity is not specific to TB only, but has also been linked to other inflammatory diseases [25]. Furthermore, propionyl-CoA is an end product of valine and isoleucine catabolism, and is a microbial metabolite, as discussed later.
Four of the eight significant metabolites – 3HPA, 5HIAA, 5HHA, and PAG – can also be linked to urinary markers of pulmonary TB as part of the altered host metabolome, caused by M. tb [26]. 3HPA is a microbial product of glycerol [27], a fatty acid that was identified as increased in TB cases by Luier and Loots [26]. 5HHA (a fatty acid) and 5HIAA (a tryptophan catabolite) were found to be increased in our study and also by Luier and Loots [26]. PAG, increased in this study, is a biotransformation product of phenylacetic acid, and also found to be increased by Luier and Loots [26].
Fig. 5
Branched-chain amino acid (BCAA) catabolism (in the host: red; and as co-metabolism (both host and microbial): purple). Elevated 3-methylcrotonylglycine (3MCG), isobutyrylglycine (IG), and 2-methylbutyrylglycine (2MBG) are urinary markers of dysbiosis at T1 of TBM treatment. It should be noted that the three BCAA intermediates (3-methylbutaonyl-CoA, isobutyrl-CoA and 2-methylbutanoly-CoA) can also be linked to branched-chain fatty acids that are specific to deterioration of the unique cell wall of M. tb (microbial specific: blue). Propionyl-CoA is also an end product that can be linked to Fig. 6
3-Hydroxypropionic acid (3HPA) – host immune response, gut microbiome and M. tb metabolismThe metabolite 3HPA was the only metabolite that was significantly (adjusted p < 0.01) increased in all three TBM stages at T1 (Fig. 3). Treatment of TBM stages 1 and 3 (Fig. 4) showed 3HPA returning to that of the control levels by T6. TBM stage 2 showed a similar linear trend as stages 1 and 3, up to T4; thereafter, there was a re-emergence of increased 3HPA. Elevated levels of 3HPA in the host usually can be caused by defect in the enzyme propionyl-CoA carboxylase. Certain metabolic conditions in the host have been linked to appearance of high concentration of 3HPA in urine. These include: propionyl-CoA carboxylase deficiency, combined carboxylase deficiencies resulting from a defect of biotin metabolism, methylmalonic semialdehyde dehydrogenase deficiency, and 4-hydroxybutyric aciduria [28,29,30,31]. Significant amounts of 3HPA can bring about functioning as an acidogen (induce metabolic acidiosis) and metabotoxin (prompts adverse effect at chronic high levels) [32, 33]. In a study reported by Pollitt et al. [29], three infants died due to a peculiar pattern of a significant urinary excretion of 3HPA, attributed to abnormal bacterial metabolism in the gut. 3HPA is an intermediary in the degradation of gut-produced propionic acid and branched-chain amino acids, as well as an intermediary metabolite in the microbial bi-cycle of propionyl-CoA and acetyl-CoA, and for carbon fixation via the microbial cycle of hydroxypropionic acid and hydroxybutyric acid (Fig. 6). Furthermore, 3HPA is a key intermediate in propionyl-CoA metabolism, which is used by M. tb via three mechanisms: (1) the protein ISL1 converts propionyl-CoA to succinic acid and pyruvic acid for energy production via either isocitrate lyase in the glyoxylate shunt or methylisocitrate lyase in the methylcitric acid cycle; (2) propionyl-CoA carboxylase converts propionyl-CoA to methylmalonyl-CoA for the vitamin B12-depedent methylmalonyl pathway, which is used for for succinyl-Coa production; (3) propionyl-CoA, in the form of methylmalonyl-CoA, is used to generate building blocks for the M. tb cell wall [34]. Hence, 3HPA is a metabolite of the host, but also has strong association with altered gut microbiota metabolism; our study and previous reports indicate a clear link to M. tb metabolism.
Fig. 6
Microbial-specific (blue background) bi-cycle of acetyl-CoA, propionyl-CoA and carbon fixation via the microbial cycle of hydroxypropionic acid and hydroxybutyric acid. Both these microbial cycles rely upon the intermediary 3HPA, which is elevated and associated with dysbiosis during TBM.
Prolonged immune response-related protein catabolism to treatment and induced M. tb deathIG, 3MCG and 2MBG were significantly elevated at T1 in TBM stages 2 and 3 (Fig. 3). In Fig. 4 it can be seen that for TBM stage 1, both 3MCG and 2MBG remained above that of the controls at a constant level (i.e., no trend), whereas IG for TBM stage 1 inexplicably increased during the course of treatment. For all three of these metabolites (IG, 3MCG and 2MBG), there was a clear linear trend of being significantly elevated at T1 and returning to control concentration levels by T6, for TBM stage 2 and 3. Loots et al. [35] also reported elevated urinary concentrations of 2MBG, and other organic acids associated with a multiple acyl-CoA dehydrogenase defect (MADD) metabolic profile in the urine of TB patients, supporting the result in this study. Chronic infection has been shown to cause vulnerability of the host, thus creating a cycle for survival, thereby activating specific metabolic pathways resulting in hypermetabolism, increased nitrogen loss, and enhanced gluconeogenesis. M. tb largely depends on human-derived nutrients for survival and replication including a source of nitrogen. Amino acids, including the BCAA amino acids isoleucine, leucine, and valine, are the most important organic nitrogen suppliers required by M. tb for intracellular growth, and survival against intracellular host-induced stress. All of the above contributes to TB cachexia, and subsequently, the WHO recommends, a patient with TB should consume more protein than is suggested for a healthy person (an additional 15–30% protein) [36,37,38,39,40]. Furthermore, 3MCG, IG and 2MBG can also be considered end products of β-oxidation of methyl branched-chain fatty acids of the M. tb cell wall, and the changes seen in our study can also be associated with M. tb death, or M. tb load.
Differences in tryptophan metabolism between disease severity and time5HIAA, a microbial catabolite of tryptophan, was significantly elevated in TBM stages 2 and 3 at T1, and returned close to control levels after 6 months of treatment. However, 5HIAA at TBM stage 1 remained elevated at a constant level throughout treatment. The tryptophan pathway has been indicated in studies as a key metabolic pathway in TBM [41, 42]. Tryptophan, a precursor used to synthesize the biogenic amine serotonin (5-hydroxytryptamine), is the main signalling molecule involved in several physiological processes and controls gut motility [43,44,45]. Serotonin, biosynthesized from tryptophan, functions as a neuromodulator and maintains homeostasis as a neuroendocrine and neurotransmitter [46]. Urinary excretion of 5HIAA serves as a marker for determining serotonin in neurological conditions, including Friedreich’s ataxia, schizophrenia, and olivo-ponto-cerebral atrophy [46,47,48]. The presence of 5HIAA in urine has been reported in a patient with carcinoid heart disease (CHD), with a high significant level as compared to patients without CHD. Kaltsas et al. [49] further confirmed that higher urine 5-HIAA levels are linked to an increased risk of developing CHD. Cheetham et al. [50] reported low levels of 5HIAA in cerebrospinal fluid of patients with depression, indicating increased serotonin breakdown. The biosynthesis of serotonin occurs in serotonergic neurons within the central nervous system in minor quantity, as well as in the enterochromaffin cells that lie along the gastrointestinal tract, where a majority is synthesized. 5HIAA is also a derivative of microbial indole-3-acetic acid and has been found to be a product of human gut microbiota, mostly synthesized in the kidney and liver, and passed in the urine [51, 52]. The microbe-produced indole derivatives of tryptophan have been shown to have a clear link to immune regulation in the host [53]. 5-Hydroxyindole (5HI), a microbial product and possible precursor of 5HIAA, is known to be involved in CNS regulation and antioxidant activity. 5HI protects cells by attenuating oxidative stress and consequently protects against mitochondrial dysfunction [54]. Liu et al. [55] shows that the microbial tryptophan metabolite 5HIAA can be used to assess intestinal homeostasis. Enrichment of microbial 5HIAA activates the Epac/Rap1 signalling pathway, which plays an important role in intestinal homeostasis Zhang et al. [56]. Furthermore, multi-omic profiling has shown that 5HIAA is a sensitive marker for intra-abdominal hypertension and sepsis in the host, and also that tryptophan-related metabolites (especially 5HIAA) are associated with microbiota changes, with several microbiota species correlating (positively and negatively) with 5HIAA [57]. Thus, 5HIAA is clearly a host metabolite, but is also a microbial metabolite, making it a co-metabolite. However, what is not clear from the literature is how 5HIAA is produced by gut microbiota. Moreover, a key question we ask is this: if increased flux of tryptophan into the kynurenine pathway is known to occur due to upregulated IDO-1 activity during TB, and other diseases [53, 58], then where/how is there sufficient bioavailability of tryptophan in the host for the increased levels of urinary 5HIAA that have been observed in this study?
Disturbances in fatty acid metabolismIn our study, (5HHA) was significantly increased in TBM stages 2 and 3 at T1 and showed a decreasing linear trend that approached control levels for all three TBM stages by T6. 5HHA is a fatty acid dicarboxylic acid breakdown product that has been identified as a urinary microbial metabolite, under situations of fasting, aberrant fatty acid oxidation (FAO), and medium-chain triglyceride (MCT) feeding [59,60,61]. With the aim of elucidating mechanisms of TB, Luier and Loots [26] found anomalies in the host’s fatty acid and amino acid metabolism brought on by pulmonary TB. These fatty acids and degradation products comprised 5HHA, glycerol monostearate and 2-octenoic acid. Note that 2-octenedioic acid was significantly elevated in this study at T1, T3, T4, T5 and T6. Other fatty acids that arose at later treatment times as significant in this study were pimelic acid and suberic acid. It is known that M. tb uses fatty acids as primary sources of energy results; hence increased urinary fatty acids and their catabolic products are common in TB cases. Additionally, other factors that might influence the host’s response are different bacterial processes, which could help to elucidate the associated loss of weight in TB cases [26, 62, 63]. Hence, disturbances in fatty acid metabolism are another landmark of gut dysbiosis induced by TB. However, to our knowledge, 5HAA has not been directly linked to changes in gut microbiota.
Perturbed tyrosine–phenylalanine metabolismLastly, 4HHA was significantly increased in TBM stages 2 and 3 at T1, and PAG was significantly increased only in TBM stage 1 at T1. Both metabolites showed similar linear trends – TBM stage 1 stayed elevated throughout treatment, whereas TBM stages 2 and 3 showed a decreasing trend, approaching control levels by T6. 4HHA and PA are components of host tyrosine–phenylalanine (aromatic amino acids) metabolism, and metabolites of microbial origin. 4HHA and PA have been identified as a major consequence of perturbed metabolism associated with diseases linked with alterations to the gut-microbiome [8, 40, 64,65,66]. A study by Mason et al. [8] examined the urinary profiles of (untreated) infants and children (≤ 13 years) from cohorts of 12 confirmed TBM cases, 12 suspected TBM cases that were later confirmed negative for TBM, and 29 healthy controls. Mason et al. [8] identified four metabolites, including 4HHA, as having potential for a non-invasive diagnostic biosignature, as well as an increase in PAG. In another study, Das et al. [67] compared the urinary metabolic profiles of patients with active pulmonary TB (n = 11) under treatment to those of healthy controls (n = 11). The urine metabolome of follow-up samples revealed a treatment-dependent trend for numerous metabolites, notably 4HHA; those who were deemed clinically healed displayed a metabolic profile comparable to that of asymptomatic healthy individuals, suggesting that a possible target for TB diagnosis and treatment is revealed by disrupted tyrosine–phenylalanine metabolism [67]. Children with autism have also been found to produce abnormal PA excretion through the urine [68]. PA has also been linked to liver dysfunction and this aligns with the pathogenesis of TBM; namely, by TBM stage 3 the patient has had the infection for two or more weeks and the accumulated hepatotoxins have been responsible for some damage to the liver. This could explain why only TBM stage 3 cases showed significantly increase PA in our study.
Urinary biomarkers for M. tbOf note, methylcitric acid (MCA) was identified as significant at T3, T4 and T5, and methylsuccinic acid (MSA) was observed as significant at T5 and T6. MCA and MSA have been linked to M. tb metabolism by several studies [8, 69,70,71,72]. Therefore, we postulate that some of the TBM patients retained a reservoir of M. tb somewhere in their body – that is, M. tb was not entirely eradicated by treatment. This is an untested postulate, but one that we would like to test in future treatment studies.
Limitations of this studyPrevious work has shown that tuberculostatic antibiotics cause changes in the gut microbiota community, and associated metabolism [73, 74]. These findings have not escaped our attention. It should be stated that we cannot definitively demarcate which metabolites are strictly the response of treatment or disease. Treatment invariably has some effect(s). For our study, however, we focused on eight altered metabolites that we consider to be most closely associated with the disease – TBM, and its associated dysbiosis. Other limitations of this study are that follow-up M. tb tests were not done and there are no data on the status of the TBM patients after treatment. Finally, the time of baseline sample (T1), collected upon release from hospital, was variably different for each TBM patient (an uncontrolled confounder). Each TBM patient therefore required a unique amount of time to stabilize in hospital before being discharged.
留言 (0)