
記住我


Familial exudative vitreoretinopathy (FEVR), as described for the first time in 1969 by Schepens and Criswick, is a hereditary condition which is characterized by peripheral retinal abnormalities and incomplete vascularization [8]. The presence of incomplete and aberrant vascularization is associated with the development of a variety of eye complications, which includes retinal exudates and neovascularization, retinal folds and detachments, vitreous hemorrhage, macular ectopia, and eventually resulting in total blindness.
FEVR has great phenotypic and genetic heterogeneity. Inheritance of FEVR, which is genetically determined, can be dominant, recessive or X-linked, with dominant inheritance being the most common manner. It has been currently found that at least nine genes are identified as responsible for the development of FEVR, which includes NDP, FZD4, LRP5, TSPAN12, ZNF408, KIF11, RCBTB1, CTNNB1, and JAG1 [9]. The proteins from the initial four genes work together in the Norrin/β-catenin signaling pathway, known as the Norrin/Frizzled-4 signaling pathway as well, and exhibit strong mutual interaction [10].
In previous study, we characterized the subtypes of FEVR by genetic analyses combined with clinical symptoms. Significant phenotypic difference was shown in TSPAN12 mutations with ocular developmental disorder affecting retinal exudates and neovascularization, retinal folds and detachments, vitreous hemorrhage, and macular ectopia [11]. In this study, we investigated a cohort of FEVR patients with TSPAN12 mutations. Compound heterozygous or homozygous TSPAN12 variants were found in 6 affected patients with a unique combination of lens and iris abnormalities.
Approximately 47.87% (56 of 117) of the variants were found in 7 genes (FZD4, LRP5, TSPAN12, NDP, CTNNB1, ZNF408, and KIF11) among the variants recognized in these study participants. 9 probands were identified as carrying TPSAN12 mutation, with a mutation detection rate of 15.25%. The highest non-missense mutation proportion was TSPAN12, reaching 44.44%.
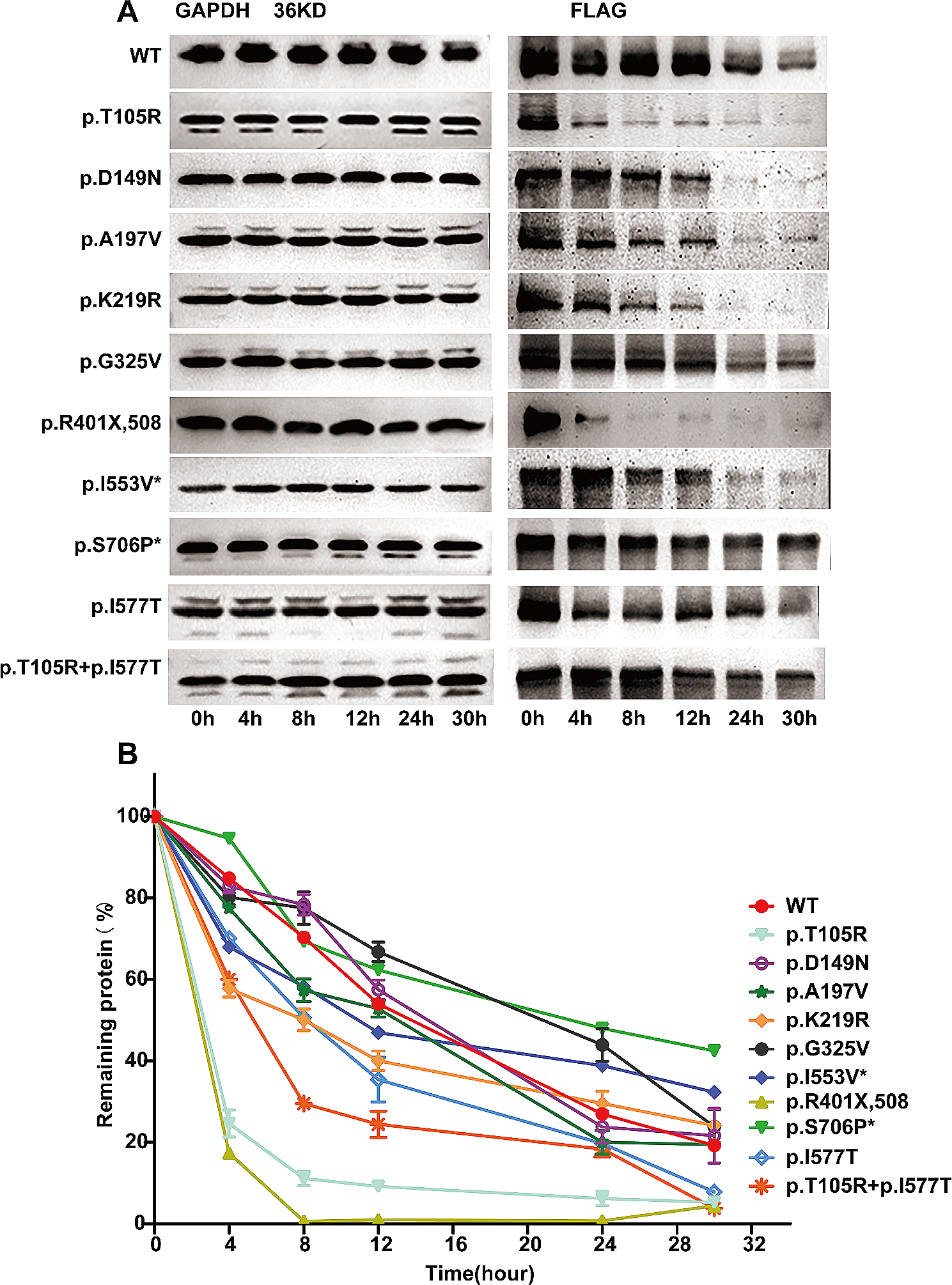
In this study, these TSPAN12 variants were identified as being deleterious by virtue of the results of bioinformatic analyses and the rareness in general population, but further functional experimental research were needed to formally confirm their deleterious properties. To understand the clinical significance of TPSAN12 variants, functional study of the 3 mutations (c.612+1G>A, c.149+2 T>C and c.360G>A) on TPSAN12 expression was made in this study. Minigene analysis confirmed the 3 mutation on TPSAN12 found in FEVR patients caused alters splicing resulting in loss of TPSAN12 expression.
The novel intronic variants in TPSAN12 was shown a deleterious effect on silico algorithms. An analysis of mini-gene splicing assay in vitro revealed that these variants alter splicing, causing frameshifts and predicted premature termination codons, resulting in protein dysfunction from this allele via nonsense-mediated mRNA decay (NMD).
TSPAN12 is a tetraspanin which is encoded by chromosome 7q31 and composed of 305 amino acids. An intracellular loop and two extracellular loops (ECL-1 and ECL-2) comprise four transmembrane domains. TSPAN12 was revealed to associate selectively with Norrin/β-catenin pathway as the significant auxiliary constituent part of Norrin/FZD4/LRP5 complex [12]. Through interacting between the FZD4 and LEL, TPSAN12 is anchored into the Norrin receptor complex. It improves its signal transduction and enhances the selectivity of ligand-receptor binding by promoting the polymerization of Norrin/FZD4/LRP5 complex, and ultimately assists in reducing the phosphorylation and degradation of β-catenin, making more β-catenin into the nucleus to act as transcription factors. TSPAN12 participates in retinal angiogenesis significantly under physiological conditions through Norrin/β-catenin signaling pathway. In contrast, TSPAN12 mutations have been reported to be associated with vascular injury, nerve cell damage, microaneurysms and other retinal vascular diseases [13].
In patients who have complex heterozygous mutations, inactivation of the PTC-containing allele by NMD results in a halving of the total amount of effective TSPAN12 proteins, thus possibly becoming the reason of diseases due to haploinsufficiency. The dominant system between Norrin/β-catenin pathway and Wnt/β-catenin pathway systems presents a difference between numerous regions of the central nervous system, among which the Norrin/β-catenin pathway system makes the major contribution to the retina [14]. Recessive loss-of-function mutations in TPSAN12 results in aberrant Norrin/β-catenin pathway, rather than affecting Wnt/β-catenin pathway. Therefore, the imbalanced signaling and relative enhancement of the latter become potential risk factors of diseases associated with TPSAN12 and FEVR. The specific promotion of the Norrin/β-catenin pathway owing to TSPAN12 mutations will cause spectrum of retinal vascular diseases (Fig. 7).
Fig. 7
The proposed mechanisms of how TSPAN12 mutations lead to FEVR
The detailed molecular mechanism by TSPAN12 mutations is still lacking. Further integrative functional studies in the comprehension to TSPAN12 sequence variants is needed. For gaining a deeper understanding of the influence of TSPAN12 in ocular system, animal model is needed to build in our further study.
留言 (0)