
記住我


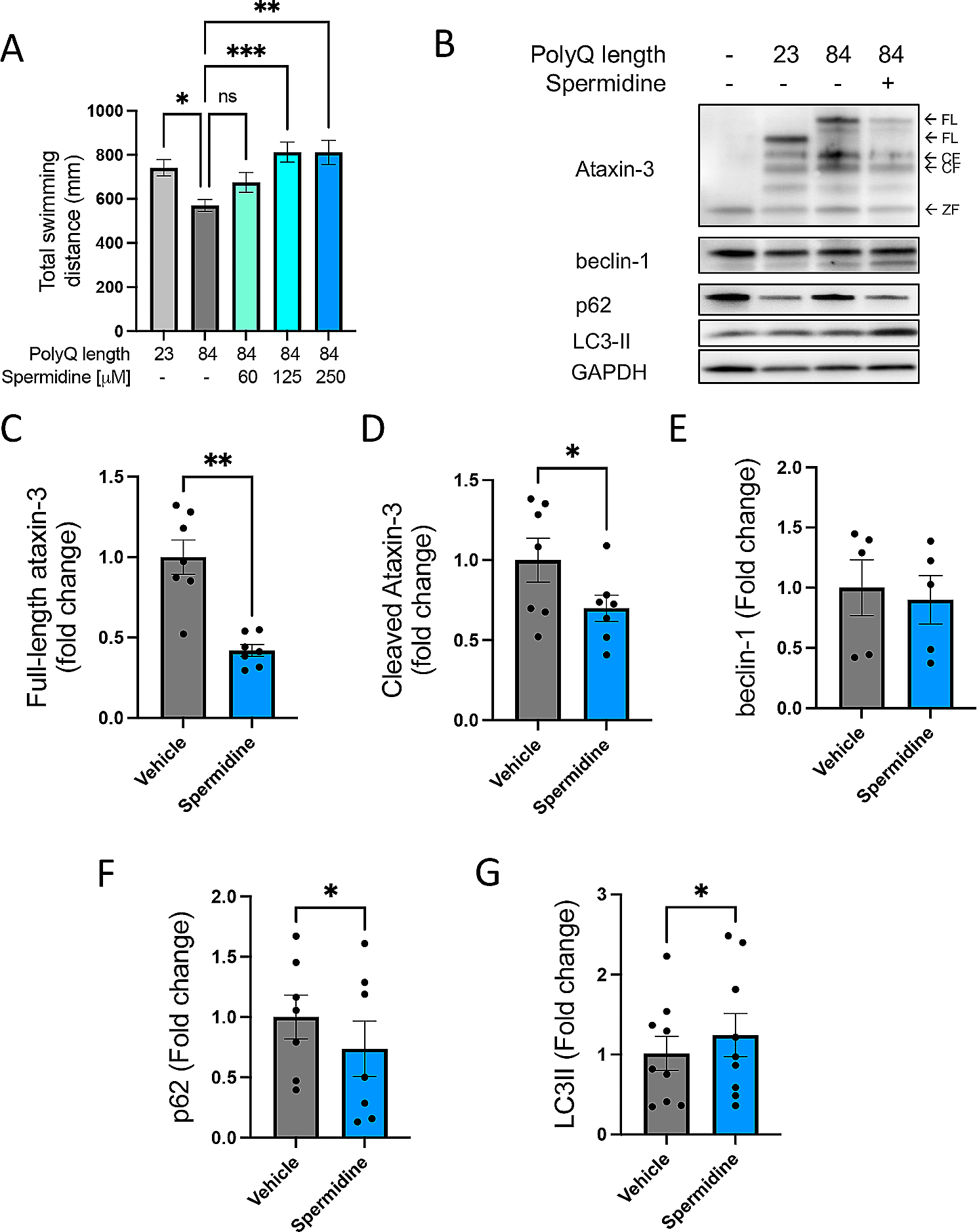


Burst firing of dentate GCs has been observed both in vivo [5] and ex vivo [7], but its physiological significance in synaptic plasticity is not well understood. We examined firing patterns of mature GCs that have input resistance (Rin) less than 200 MΩ in response to somatic current injection [15,16,17,18,19]. When we applied a step current just above action potential (AP) threshold (rheobase current) for 1 s in whole-cell current clamp mode, a group of cells generated APs in bursts, doublet in majority (82.8%, 18 of 22) and sometimes triplet (18.2%, 4 of 22), while others showed regularly spiking patterns (Fig. 1A). The histogram of initial firing frequency (Finit) showed bimodal distribution (Fig. 1B), so that we nominated cells with Finit under 50 Hz as regular-spiking (RS, black), while cells with Finit over 50 Hz as burst-spiking (BS, red) neurons. The mean value for Finit was 10.6 ± 2.2 Hz (n = 18) in RS-GCs and 147.1 ± 11.2 Hz (n = 22) in BS-GCs. As the injection current increased, Finit increased in RS-GCs, and the difference of Finit between RS- and BS-GCs gradually disappeared (Fig. 1Cc). Despite the remarkable difference in Finit, the number of APs during 1 s depolarization was not significantly different between two groups (Fig. 1Cb). Analyses of AP shapes revealed that the threshold voltage for AP generation was lower, AP duration was longer, and afterhyperpolarization (AHP) was smaller in BS-GCs compared to those in RS-GCs (Fig. 1D). No significant difference was found in passive electrical properties such as input resistance (Rin) and resting membrane potential (RMP) (Fig. 1E). Interestingly, in DG-GCs that have Rin more than 200 MΩ, which are less mature according to the criteria of maturation [20, 21], bursting was very rarely observed (Fig. 1F), suggesting that burst firing is a characteristic feature of fully mature DG-GCs.
Fig. 1
Intrinsic properties of regular-spiking (RS) and burst-spiking (BS) mature granule cells. A Representative voltage responses of RS (black) and BS (red) cells to somatic current injection of 250, 300 and 650 pA (1 s duration). Inset, initial firing of BS-GCs at expanded time scale. Initial firing frequency (Finit) was measured as the frequency of first two APs at rheobase current injection. B Bimodal distribution of Finit among mature GCs. Mature GCs were divided into RS and BS with the reference frequency of 50 Hz (RS/BS, n = 18/42). C, a, b Finit (a) and spike numbers (b) of RS and BS cells as a function of injected current amplitude (from 50 to 650 pA). Finit of RS cells increased steeply compared to that of BS-GCs. The general excitability of both GCs was not different throughout all steps (RS/BS, n = 18/22). D Summary bar graphs for analyses of 1st AP waveform evoked by somatic rheobase current injection into RS and BS cells. Mean values for AP threshold were − 34.9 ± 0.5 mV in RS and − 36.7 ± 0.7 mV in BS (*p < 0.05). For afterhyperpolarization (AHP) amplitudes, 17.1 ± 0.5 mV in RS and 12.0 ± 0.5 mV in BS (***p < 0.001). For half-width duration, 0.70 ± 0.01 ms in RS and 0.79 ± 0.01 ms in BS (***p < 0.001; RS/BS, n = 18/22). E Input resistance (Rin; RS, 116.1 ± 9.6 MΩ; BS, 115.3 ± 6.1 MΩ) and resting membrane potential (RMP; RS, − 82.1 ± 1.3 mV; BS, − 81.2 ± 0.9 mV) were not different between RS and BS. F Proportion of RS and BS cells depends on the GC maturity. BS cells were more frequently found in the group of mature GCs (Rin < 200 MΩ) compared to the less mature GC group (Rin ≥ 200 MΩ; RS/BS, n = 16/17). G Distributions (left) and mean values (right) for maximal dendritic branch order in RS (black) and BS (red) cells (RS, 4.5 ± 0.3, n = 8; BS, 5.1 ± 0.1, n = 18, p = 0.10). H Proportion of RS and BS cells along the dorsoventral axis (RS/BS in dorsal, n = 84/148; in ventral, n = 37/8). Error bars indicate S.E.M. *p < 0.05. ***p < 0.001. n.s., not significant (p > 0.05)
To explore whether the bursting behavior is related to morphological properties of GCs, we counted the maximal branch order (MBO) from z-sections of confocal images of biocytin-filled RS- and BS-GCs (Additional file 1: Fig. S1). The MBO of majority (65.4%) of mature GCs was five, while cells with MBO higher than five was only found in BS-GCs and that with lower than 4 is only found in RS-GCs (Fig. 1G). The average MBO of BS-GCs, however, was not different from that of RS-GCs (RS, 4.5 ± 0.3, n = 8; BS, 5.1 ± 0.1, n = 18, p = 0.10, Mann–Whitney test). We then examined whether the relative proportion of BS- and RS-GCs differs along the hippocampal dorso-ventral axis. We found that BS-GCs were dominant in the dorsal DG, and its proportion was opposite in the ventral DG. Among 232 recorded neurons in dorsal DG, 148 (64%) GCs were identified as BS-GCs, while only 8 (18%) out of 45 GCs were BS-GCs in ventral DG, indicating that the dorsal DG harbors more BS-GCs compared to the ventral DG (Fig. 1H).
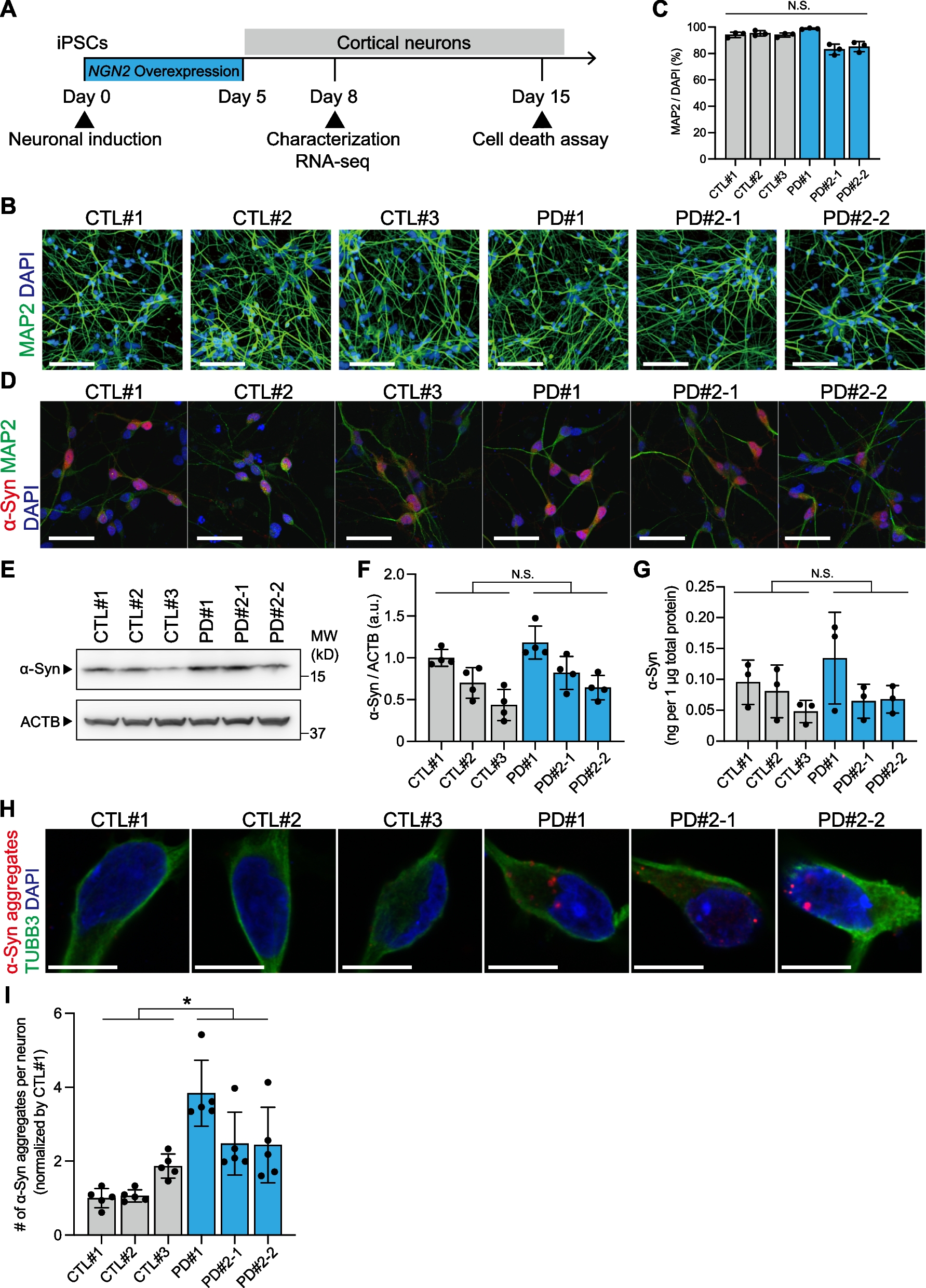
Subthreshold EPSP summation evoked by a single bout of HFS induces NMDA receptor-dependent LTP at LPP-GC synapsesTo investigate whether intrinsic firing patterns have any effects on long-term synaptic plasticity, we recorded excitatory postsynaptic potentials (EPSPs) from RS-GCs or BS-GCs by stimulating lateral perforant pathways (LPP) in the presence of PTX (100 μM, a GABAAR blocker) and CGP52432 (1 μM, a GABABR blocker) (Fig. 2A). After measuring the baseline EPSPs evoked by stimulation of LPP in a 10 s interval for about 5 min, a single bout of high frequency stimulation (HFS, 10 stimuli at 100 Hz) was applied. For the HFS, we tested two different levels of electrical stimulation intensity: low intensity to induce subthreshold response (HFSL) and high intensity to evoke at least 3 APs (HFSH). Compared to MPP-EPSPs, LPP-EPSPs exhibited significantly higher paired pulse ratio and larger normalized area of HFSL-evoked EPSP summation to the 1st EPSP amplitude (Additional file 1: Fig. S3A; Figs. 2B vs. 7B). The average stimulation intensities for HFSL and HFSH of LPP were 15.6 ± 0.9 V (n = 21) and 25.7 ± 1.4 V (n = 18), respectively (Additional file 1: Fig. S3B). The average amplitudes of baseline EPSPs induced by HFSL and HFSH were 5.9 ± 0.3 and 13.8 ± 1.0 mV, respectively (Additional file 1: Fig. S3C). Temporal summations of EPSPs evoked by HFSL reached their peaks between − 60 mV and − 40 mV at the 6th or 7th stimulus. RS- and BS-GCs showed no detectable difference in the temporal summation kinetics (RS, black; BS, red; Fig. 2B). Despite that HFSL of LPP evoked no postsynaptic AP, it induced long-term potentiation (LTP) of EPSP amplitudes in both GC groups, which lasted at least 30 min (Fig. 2C). We denoted this form of LTP as LTPsub, which stands for LTP induced by subthreshold stimulation. The increase in baseline EPSP amplitudes after HFSL was not different between BS-GCs (36.9 ± 8.9%, n = 13) and RS-GCs (35.2 ± 5.3%, n = 12, p = 0.65). The magnitude of LTPsub was correlated with the peak of EPSP summation (r = 0.54, p < 0.001), and significant LTPsub was induced when the peak was higher than − 60 mV (Additional file 1: Fig. S3D). To examine the involvement of NMDA receptor (NMDAR) in LTPsub, we tested the effect of APV (50 μM, a NMDAR blocker) on EPSP responses and LTP expression induced by HFSL. APV profoundly suppressed the baseline EPSPs as well as EPSP summation (Fig. 2E), and abolished LTPsub (Fig. 2F). These results suggest that NMDAR-dependent LTP can be induced at LPP-GC synapses by a single bout of HFS that evokes only a subthreshold voltage response.
Fig. 2
NMDAR-dependent LTP at LPP-GC synapses is induced by a single bout of high frequency stimulation (HFS) at subthreshold level. A Schematic diagram illustrating the recording configuration for synaptic stimulation and whole cell recording of mature GC. Lateral perforant pathway (LPP) in outer molecular layer (OML) was electrically stimulated by a bout of HFS (10 stimuli at 100 Hz). Scale bar is 100 μm. B HFSL is defined as low intensity HFS that induce subthreshold response. HFSL-evoked subthreshold responses of RS- (black) and BS-GCs (red) (left), and their cumulative EPSP amplitudes (right). C Time courses of normalized EPSP amplitude before and after HFSL (same as in B). Each point represents averaged value for adjacent 3 EPSP amplitudes (30 s binned). Black dashed line denotes baseline EPSP. Inset, Representative traces for average of 30 EPSP traces before (a) and 26–30 min (b) after HFSL (This holds for inset traces in all subsequent figures except in Fig. 2F and Fig. 7C). D LTP magnitudes before and after HFSL. There was no significant difference between RS and BS (RS/BS, n = 12/13). E Left, Representative traces for EPSP summation in control (black) and after application of APV (brown, 50 μM). Right, Mean values for EPSP area in RS [3.0 ± 0.3 mV s (Con) vs. 1.3 ± 0.2 mV s (APV), n = 5, *p < 0.05] and in BS [2.9 ± 0.3 mV s (Con) vs. 1.9 ± 0.1 mV s (APV), n = 5, *p < 0.05]. Note that the APV effect on subthreshold EPSP summation was examined at synapses which have already underwent LTPsub. F Time course of normalized EPSP before and after HFSL in the presence of APV in both GCs. EPSP amplitude was not potentiated (RS, 1.7 ± 6.0%, n = 3, light brown; BS, − 2.5 ± 4.5%, n = 4, brown). Inset, EPSPs averaged over 1 to 5 min before (a) and after (b) HFSL. Shades and error bars, S.E.M. *p < 0.05. n.s., not significant (p > 0.05)
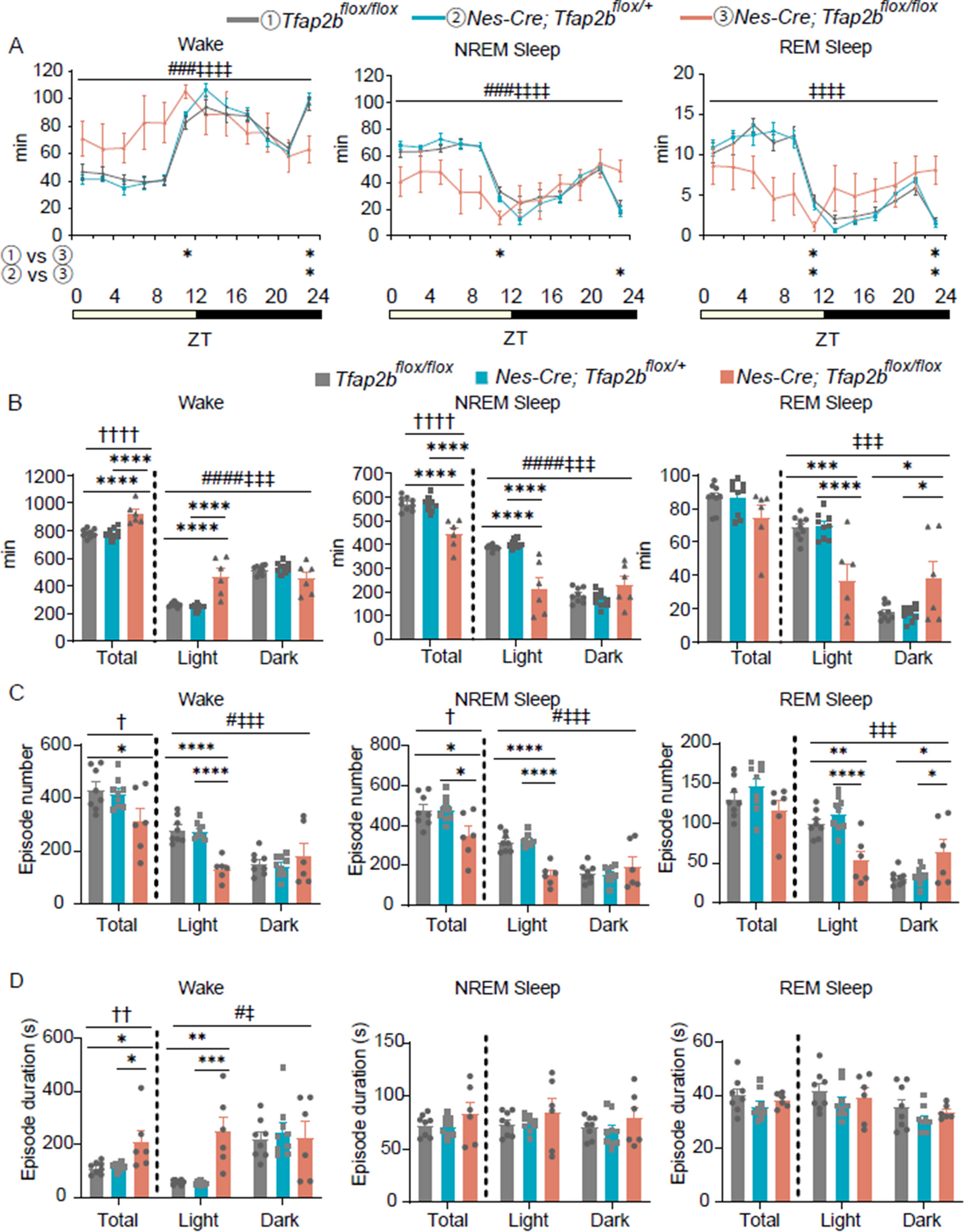
Postsynaptic burst firing is essential for Hebbian LTP at LPP-GC synapsesWe then examined whether AP firings in response to HFSH show any difference between BS and RS (Fig. 3A). The Finit of HFSH-evoked APs was mostly higher than 100 Hz in BS-GCs (128.3 ± 6.9 Hz, n = 21, Fig. 3A). Furthermore, BS-GCs showed a moderate correlation between the Finit of synaptically evoked APs and that of APs evoked by somatic stimulation (r = 0.55, Fig. 3B). In contrast, the Finit of HFSH-evoked APs in RS-GCs was significantly lower than that in BS-GCs (92.0 ± 9.9 Hz, n = 18; p < 0.01; Fig. 3A). These results suggest that mechanisms underlying intrinsic firing pattern contribute to synaptically evoked firing pattern. When the 2nd HFS with higher stimulation intensity (denoted as ‘HFSH-2’) was applied 10 min after HFSL by which LTPsub has been already expressed both in RS and BS, HFSH-2 induced further potentiation of EPSPs in BS-GCs, but not in RS-GCs (Fig. 3C). The time course of this LTP induced by HFSH-2 is shown as the EPSP amplitudes normalized to the EPSP amplitude just before applying HFSH-2 (Fig. 3D). The increase in the EPSP amplitude at 30 min was 44.0 ± 4.8% (n = 7) in BS-GCs, but negligible in RS-GCs (− 4.2 ± 7.0%, n = 12; p < 0.001). These results indicate that BS-GCs express postsynaptic AP-dependent LTP (denoted as LTPAP) distinct from NMDAR-dependent LTPsub. There was a positive correlation between LTPAP magnitudes and Finit of synaptically evoked APs (Fig. 3E). Moreover, when only 1 or 2 APs were elicited by HFS with medium intensity (HFSM), LTP was not induced or not maintained even in BS-GCs (− 10.6 ± 12.8%, n = 6; Additional file 1: Fig. S4A, B), indicating that postsynaptic AP bursts comprised of at least 3 APs at the frequency higher than 100 Hz are essential for the induction of LTPAP.
Fig. 3
Hebbian LTP depends on post-synaptic AP bursts, and can be induced only in BS. A Left, Representative voltage responses in RS (black) and BS (red) to HFSH-2 which elicited 3 APs. HFSH-2 is defined as high intensity HFS that was applied following HFSL. Right upper, The boxed traces are superimposed for comparison at expanded time scale. Right lower, Initial AP frequency (Finit) of each group (RS/BS, n = 18/21; **p < 0.01). B Relationship between Finit of APs evoked by somatic current injection and that by synaptic stimulation. Two parameters were significantly correlated in BS (r = 0.55, **p < 0.01) but not in RS (r = 0.11). Black bold line, linear regression in BS. r, Pearson’s correlation coefficient. C Time course of normalized EPSP changes induced by applying two sequential HFS (HFSL and HFSH) in RS (black, n = 5) and BS (red, n = 5). Note that LTPAP was induced on top of LTPsub in BS, not in RS. D Left, Time course before and after HFSH-2. Right, Magnitude of LTPAP in RS- and BS-GCs (RS: − 4.2 ± 7.0%, n = 12; BS, 44.0 ± 4.8%, n = 7, ***p < 0.001). E LTP magnitude as a function of synaptically evoked Finit. Finit was correlated to LTP magnitude (r = 0.50, *p < 0.05). Open circles, individual data; closed circles, averaged value for each group. Black line, linear regression line. F Pairing protocol for LTPAP induction. It consists of subthreshold HFS (HFSL-2) and post-synaptic 3 APs evoked by somatic pulses (2 ms, 3 nA at 100 Hz). G Left, Time course of normalized EPSP before and after a pairing protocol. Right, Pairing protocol-induced LTPAP in RS- and BS-GCs (RS/BS, n = 8/6). H LTP was not induced in the presence of intracellular solution containing BAPTA (10 mM, pink, n = 7). The pairing protocol-induced LTP time courses in RS and BS under control conditions shown in G were merged, and superimposed in gray (n = 14). I Left, Representative voltage traces evoked by HFSH-2 in the presence of APV (brown, 50 μM). No AP burst was elicited in the presence of APV. Right, Time course of normalized EPSP before and after HFSH-2. J Similar as in I, but a pairing protocol was applied instead of HFSH-2. The control trace (gray) was reproduced from panel H for comparison. K Early (LTP5, open circle) and late phase (LTP30, closed circle) LTP induced by a pairing protocol with and without APV (Control, n = 14; APV, n = 11). Shades and error bars, S.E.M. *p < 0.05. **p < 0.01. ***p < 0.001. n.s., not significant (p > 0.05)
To further test the importance of AP frequency for induction of LTPAP, we applied a pairing protocol, in which 10 EPSPs were evoked by HFSL coinciding with 3 APs at 100 Hz evoked by brief current injection to the soma (Fig. 3F, see Methods). The pairing protocol was delivered to the GCs that have already underwent subthreshold LTP. The pairing protocol successfully induced LTP regardless of cell types with no significant difference in the LTP magnitude between RS-GCs and BS-GCs (Fig. 3G), but 3 APs at 50 Hz failed to induce LTP (Additional file 1: Fig. S4C). These findings show that RS-GCs could express LTPAP as if BS-GCs did as long as high frequency APs are paired with synaptic stimulation. Therefore, we did not distinguish BS and RS but pooled the BS and RS data when we analyzed LTP response induced by pairing protocol (gray trace in Fig. 3H). LTP was not induced when intracellular Ca2+ was chelated with a high concentration of BAPTA (10 mM, Fig. 3H). These results confirm that at least 3 APs at the frequency higher than 100 Hz are required to activate LTPAP, and suggest that Ca2+-dependent mechanisms underlie this form of LTP. Finally, we examined the input specificity of LTPAP by monitoring EPSPs evoked by two stimulation electrodes placed at the outer molecular layer. To induce LTPAP using the pairing protocol, HFSL was delivered to one of two stimulation electrodes. LTPAP was induced at the synapse which underwent HFSL paired with 3 APs at 100 Hz, but not at the other synapse (Additional file 1: Fig. S2). Because LTPAP satisfied the conditions for Hebbian LTP, henceforth we will refer to LTPAP as Hebbian LTP.
NMDAR mediates the early phase LTP and facilitates EPSP summation at LPP-GC synapsesTo examine whether LTPAP shares the same Ca2+ source with NMDAR-dependent LTPsub, we tested the effect of APV. Because APV profoundly suppressed EPSP summation (Fig. 2E), in the presence of APV it was difficult to generate 3 APs even with high intensity stimulation, and thus LTPAP was not induced (Fig. 3I), indicating that NMDAR current is critical for EPSP summation to elicit high frequency AP generation. However, we could induce LTP by the pairing protocol in the presence of APV (Fig. 3J). Because the time course of LTP development was distinct from that of control pairing-induced LTP (brown vs. gray traces in Fig. 3J), we compared the LTP magnitudes in the APV conditions with the control values for the early and late phases. To this end, we measured normalized EPSP amplitudes averaged over 1 to 5 min and over 26 to 30 min after HFS, and denoted as LTP5 and LTP30, respectively. LTP5 encompasses short-term potentiation (STP) and early phase LTP [22]. LTP5 in the APV conditions was significantly lower, while LTP30 was not different compared to the corresponding control values [LTP5, 8.6 ± 5.5 vs. 25.7 ± 4.6%, p < 0.05; LTP30, 46.4 ± 12.7 vs. 52.62 ± 7.71%, p = 0.57; APV (n = 11) vs. Control (n = 14), Mann–Whitney test, Fig. 3K]. These results suggest that the contribution of NMDAR to Hebbian LTP as Ca2+ source is limited to STP and early phase LTP at LPP-GC synapses, whereas it is essential for EPSP summation and AP burst generation.
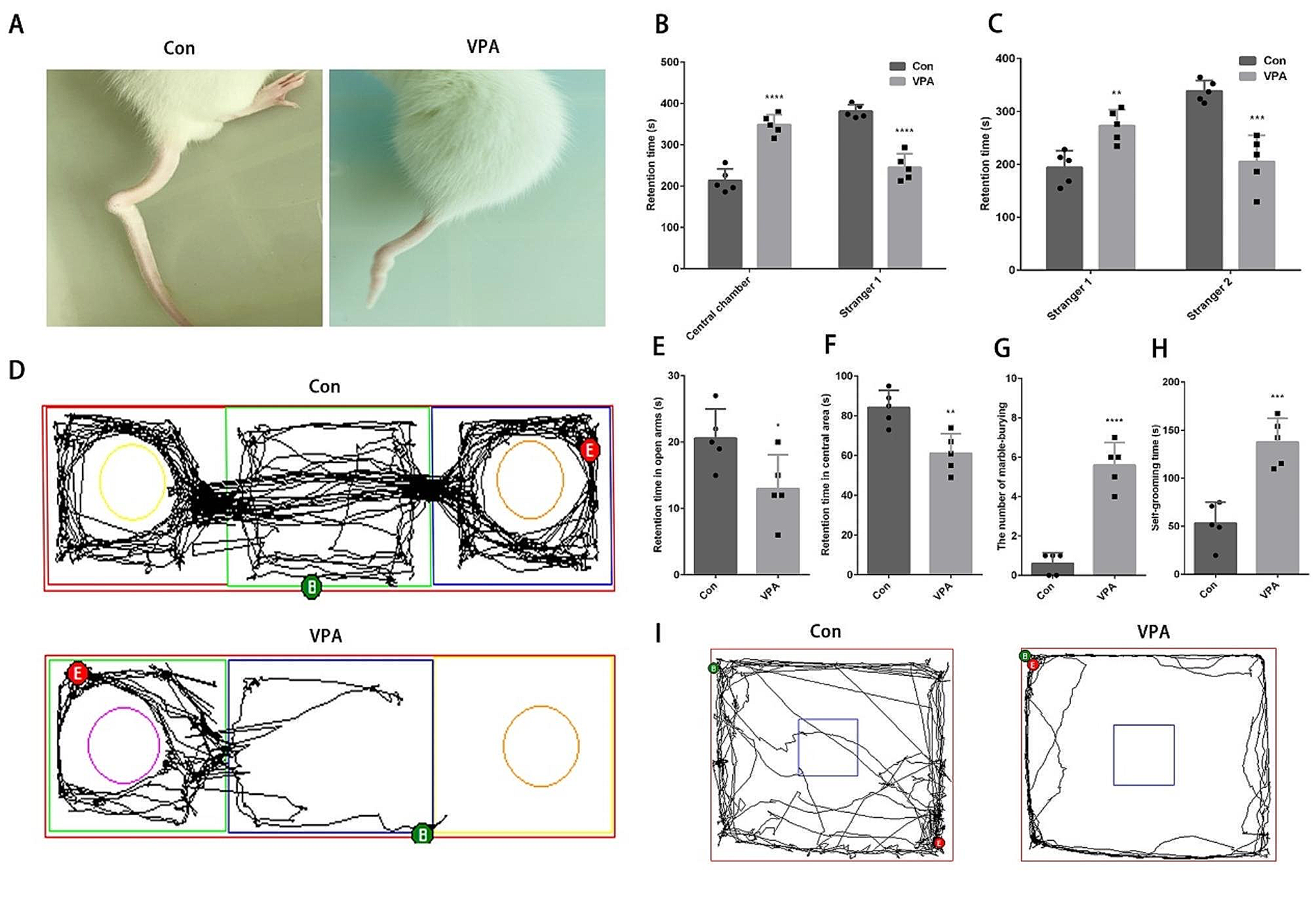
T-VDCC contributes to the late phase LTP by facilitating AP bursts at LPP-GC synapsesWe showed that burst firing evoked by somatic rheobase current injection (called intrinsic burst firing) has correlation with Finit of synaptically evoked APs which is crucial for LTPAP induction (Fig. 3). We investigated whether ion channel mechanisms underlying intrinsic burst firing also contribute to LTPAP. Since T-VDCC is known to mediate intrinsic bursting in DG-GCs [7], we investigated the role of T-VDCCs in burst firing behavior and LTPAP induction in BS-GCs. Bath application of NiCl2 (50 μM, the blocker of T-VDCC) significantly reduced Finit of intrinsic burst firing (Control, 171.8 ± 13.3 Hz; NiCl2, 38.1 ± 8.89 Hz, n = 9; p < 0.01; Wilcoxon signed-rank test; Fig. 4A). When the bursts were synaptically evoked, NiCl2 partially but significantly reduced the Finit (Control, 128.3 ± 6.9 Hz, n = 21; NiCl2, 91.2 ± 3.5 Hz, n = 13, p < 0.001, Mann–Whitney test; Fig. 4B). Nevertheless, in the presence of 50 μM Ni2+, temporal summation of EPSPs evoked by HFSL was little affected (n = 10, p = 0.11; Fig. 4C), and HFSH was able to induce LTPAP in the BS cells (Fig. 4D). In contrast to APV, the early phase LTP was preserved in the presence of Ni2+ [LTP5 of Ni2+ (n = 7) vs. control (n = 7), 24.9 ± 5.6 vs. 27.5 ± 6.3%, p = 0.90], but no further increase in the EPSP amplitudes was observed (Figs. 4D vs. 3D), and thus LTP30 was lower than the control (20.4 ± 7.7 vs. 44.6 ± 5.7%, p < 0.05, Fig. 4D). Because Ni2+ lowered the Finit of synaptically evoked AP bursts, we tested if suppression of late LTPAP can be rescued by pairing protocol. The mean value for LTP30 measured after the pairing protocol was slightly lower but not significant compared to pairing-induced LTP in control (32.6 ± 13.0 vs. 52.6 ± 7.7%, n = 8, p = 0.19; Fig. 4E), suggesting partial or little contribution of T-VDCC to the LTPAP induction. Similar to HFSH-induced LTP, LTP5 was not different from the control value (25.0 ± 8.2 vs. 25.7 ± 4.6%, p = 0.97). These results suggest that T-VDCC primarily contributes to the late phase LTPAP by enhancing Finit.
Fig. 4
T-type Ca2+ channels contributes to the late phase Hebbian LTP by facilitating AP bursts. A Left & Middle, Voltage responses of a BS cell to somatic rheobase current injection before (red, Con) and after bath application of NiCl2 (50 μM, green). Inset of left panel, the boxed trace in an expanded time scale. Right, Mean values for Finit before and after NiCl2 application (n = 9). B Left, Representative EPSP summation in a BS cell evoked by HFSH-2 of LPP in the presence of NiCl2. Right, Finit of AP bursts evoked by HFSH-2 of LPP in BS cells under control and NiCl2 (n = 13) conditions. C EPSP summation evoked by HFSL-2 (left) and the mean area (right) before and after applying NiCl2 (Con, 2.2 ± 0.2 mV s; NiCl2, 2.0 ± 0.2 mV s; n = 10). D Left, Time courses of normalized EPSP in BS cells before and after HFSH-2 with (green) and without (light red) NiCl2. The control time course was reproduced from Fig. 3D for comparison. Right, Magnitude of HFSH-induced LTP in the early (LTP5, open circle) and late (LTP30, closed circle) phases in BS cells (Control, n = 7; NiCl2, n = 7). E Similar as in D, but evoked by a pairing protocol (Control, n = 14, gray; NiCl2, n = 8, green). Shades and error bars, S.E.M. *p < 0.05. **p < 0.01. ***p < 0.001. n.s., not significant (p > 0.05)
Persistent Na+ current amplifies LPP-evoked EPSP summation and is essential for burst firingPreviously, it was shown that T-VDCC in axon initial segment plays a key role in intrinsic burst firing of GCs [7]. Whereas Ni2+ abolished intrinsic bursts (Fig. 4A), it partially reduced Finit of synaptically evoked bursts with little effect on EPSP summation (Fig. 4B, C), implying a possible involvement of dendritic channels in synaptically evoked AP bursts. As a candidate ion channel regulating intrinsic and synaptically evoked bursts, we examined persistent sodium current (INa,P). In CA1 pyramidal cells, INa,P amplifies subthreshold EPSPs leading to spatially tuned firing [23]. We measured Finit of intrinsic bursts in BS-GCs after applying riluzole (10 μM), a typical INa,P blocker [23,24,25]. Riluzole significantly reduced Finit of the intrinsic bursts (Fig. 5A) similar to its effect in CA1 pyramidal neurons [24]. In addition, it markedly suppressed summation of HFSL-evoked EPSPs (Fig. 5B). Due to the substantial inhibition of EPSP summation by riluzole, it was not possible to synaptically evoke AP bursts, even with very high stimulation intensity, and LTP was not induced (Fig. 5C). When 10 EPSP bursts induced by HFSL were paired with 3 APs (pairing protocol), however, the late phase LTP was completely rescued (LTP30, 43.3 ± 14.7 vs. 52.6 ± 7.7%, n = 10, p = 0.34 compared to pairing-induced LTP in control, Mann–Whitney test; Fig. 5E). The rescue of late phase LTP by the pairing protocol suggests that burst APs coincident with synaptic inputs is essential for the late LTP induction. By contrast, early phase LTP was significantly lower than the control value (LTP5, 9.6 ± 6.7 vs. 25.7 ± 4.6%, n = 10, p < 0.05), resulting in the LTP time course similar to that in the APV condition (Fig. 3J, K). This similarity may be explained by assuming that Ca2+ influx through NMDAR mediates the early phase LTP, and that INa,P contributes to NMDAR activation in distal dendrites by amplifying EPSP summation, which cannot be compensated by somatic bursts.
Fig. 5
Persistent Na+ current (INa,P) amplifies LPP-evoked EPSP summation and is essential for burst firing. A Left & Middle, AP trains in BS elicited by somatic rheobase current injection in control (red, Con) and after applying riluzole (blue, Ril, 10 μM). Right, Mean Finit before and after application of riluzole (Con, 176.5 ± 12.9 Hz; Ril, 5.7 ± 1.4 Hz; n = 10; **p < 0.01). B Left, EPSP summation evoked by HFSL-2 before and after applying riluzole. Right, Cumulative EPSP amplitudes in control and riluzole conditions (n = 24). C Representative voltage response to HFSH-2 (left) and time course of normalized EPSP before and after HFSH-2 (right, n = 5) in the presence of riluzole. D Similar as in C, but evoked by a pairing protocol. E Early (LTP5, open circle) and late phase (LTP30, closed circle) LTP evoked by a pairing protocol in control and riluzole conditions, showing that late LTPAP was rescued (Con, n = 14; Ril, n = 10). F Cartoon for focal application of riluzole (50 μM) at soma or dendrite during somatic current injection or HFSL of LPP. Scale bar is 100 μm. G Representative traces (upper) and mean Finit (lower) of intrinsic AP bursts with somatic (left) and dendritic (right) puff of riluzole [Soma, 128.8 ± 12.8 Hz (Con) vs. 9.0 ± 3.0 Hz (Ril), n = 12, **p < 0.01; Dend, 137.5 ± 13.3 Hz (Con) vs. 137.6 ± 14.1 Hz, n = 6]. H Similar as in G, but area of subthreshold EPSP summation evoked by HFSL-2. The EPSP summation was not reduced by somatic puff (left) but by dendritic puff (right) [Soma, 2.6 ± 0.3 mV s (Con) vs. 2.5 ± 0.4 mV s (Ril), n = 7; Dend, 2.9 ± 0.5 mV s (Con) vs. 1.1 ± 0.1 mV s, n = 8]. I Left, Procedure for measuring INa,P in GCs. Right, Representative current responses of RS- (black) and BS-GC (red) to a voltage ramp. J Peak INa,P amplitude in RS and BS cells (RS, 144.9 ± 15.6 pA, n = 10; BS, 245.4 ± 16.4 pA, n = 8; ***p < 0.001). Shades and error bars, S.E.M. *p < 0.05. **p < 0.01. ***p < 0.001. n.s., not significant (p > 0.05)
Since riluzole showed profound effects on both intrinsic and synaptically evoked firings, we hypothesized that intrinsic bursting behavior is mainly affected by somatic INa,P, while synaptically evoked AP firings are affected by dendritic INa,P. To test this, we examined the effect of focal puff application of riluzole (Fig. 5F). Peri-somatic puff application of riluzole (50 μM) considerably reduced the Finit of intrinsic burst firings in BS-GCs, whereas dendritic puff had no effect at all (Fig. 5G). On the contrary, the EPSP summation was profoundly diminished by dendritic puff, but not by peri-somatic puff of riluzole (Fig. 5H). These results support our hypothesis for the preferential roles of somatic and dendritic INa,P on the intrinsic and synaptically evoked firing behaviors, respectively. Since BS-GCs have higher Finit both for intrinsic and synaptically evoked APs than RS-GCs, we tested whether difference in INa,P density underlies the different bursting behavior between these two GC types. To measure INaP in the identified GC type, we first examined the AP responses to somatic rheobase current injection using the standard intracellular solution, carefully withdrew the pipette, and then re-patched the same cell again with Cs+- based pipette solution in the presence of Cd2+ (200 μM) and TEA (20 mM) in the bath solution to inhibit Ca2+ and K+ currents (Fig. 5I). We quantified INa,P in each type of neurons using a slowly rising ramp voltage command protocol from a holding potential of − 70 mV to 0 mV for 6 s in voltage-clamp configuration. In consistent with our hypothesis, the peak amplitude of INa,P in BS-GCs was significantly larger than that in RS-GCs (Fig. 5J).
L-VDCC is a major Ca2+ source for LTP induction but little contributes to firing propertiesAbove results indicate that NMDAR and T-VDCC partially contribute to LTPAP, and that INa,P is essential for intrinsic and synaptically evoked burst firings in BS-GCs. L-VDCC is known as a calcium source for NMDAR-independent slowly developing LTP induced by 200 Hz tetanic stimuli at CA3-CA1 synapses [26, 27]. We studied the role of L-VDCC in burst firing and LTPAP induction. Distinct from drugs tested above, nimodipine (10 μM), an L-VDCC blocker, had no effect on the Finit of APs evoked by somatic current injection (Fig. 6A). Moreover, nimodipine affected neither EPSP summation induced by HFSL nor the Finit of APs evoked by HFSH (Fig. 6B, C). Nevertheless, the late phase of LTPAP was abolished in the presence of nimodipine (LTP30, − 3.1 ± 11.0 vs. 44.6 ± 5.7%, p < 0.01, n = 6, Fig. 6D). Although the early LTP (LTP5) was not different, it was significantly reduced too when one outlier was ignored (LTP5, − 0.9 ± 5.7%, p < 0.05). Furthermore, LTP was not rescued by the pairing protocol (LTP30, − 2.9 ± 10.3 vs. 52.6 ± 7.7%, p < 0.01, n = 7, Fig. 6E), indicating that calcium influx through L-VDCC during AP bursts is essential to induce LTPAP. Although the LTP5 values for HFSH- and pairing-induced LTP were not lowered (HFSH, 7.2 ± 9.3%, n = 6, p = 0.10; Pairing protocol, 4.3 ± 8.2%, n = 7, p = 0.08, Mann–Whitney test), the early increase in normalized EPSP was transient, suggesting that it belongs to short-term potentiation (STP) which decayed within 3 min [22]. These results indicate that L-VDCC mediates both early and late phase LTP but not STP.
Fig. 6
Ca2+ influx through L-type Ca2+ channels mediates Hebbian LTP at LPP-GC synapses. A Left & Middle, Representative AP responses to somatic rheobase current injection before (red, Con) and after application of nimodipine (purple, Nimo, 10 μM). Right, Finit of intrinsic bursts was not affected by nimodipine (Con, 198.3 ± 26.8 Hz; Nimo, 206.7 ± 36.0 Hz; n = 5). B Exemplar traces (left) and mean areas (right) of EPSP summation evoked by HFSL-2 of LPP before and after applying nimodipine (Con, 2.1 ± 0.3 mV s; Nimo, 2.3 ± 0.4 mV s; n = 6). C Exemplar voltage response of BS-GCs (left) evoked by HFSH-2 and mean Finit (right) in the presence of nimodipine (Nimo, 154.8 ± 17.5 Hz, n = 5). Black dashed line on the bar graph, control mean Finit in BS cells (128.3 Hz). D Left, Time course of normalized EPSP in BS-GCs before and after HFSH-2. Right, Magnitudes of early (LTP5, open circle) and late phase (LTP30, closed circle) LTP in control and nimodipine conditions (Con, n = 7; Nimo, n = 6). The control LTP time course and magnitudes were reproduced from Fig. 3D (light red). E Similar as in C, D, but applied a pairing protocol instead of HFSH. The control LTP trace and magnitudes were reproduced from Fig. 3H (gray) (Con, n = 14; Nimo, n = 7). F Time course of normalized EPSP in RS- and BS-GCs before and after HFSH-1. G Left, Time course of normalized EPSP in BS-GCs before and after HFSH-1 in the presence of nimodipine. Right, Magnitudes of late phase LTP (LTP30, closed circles). Note little effect of nimodipine on HFSH-1-induced LTP in BS-GCs. Shades and error bars, S.E.M. *p < 0.05. **p < 0.01. n.s., not significant (p > 0.05)
The LTPAP so far was induced by HFSH-2 which was delivered following HFSL-1 in BS-GCs (Fig. 3C). We wondered whether L-VDCC would contribute to LTPAP induced by HFSH-1 (HFSH without preceding HFSL) to a similar extent as HFSH-2-induced LTPAP. The LTP magnitude induced by HFSH-1 was significantly less than the sum of LTP magnitude induced by the HFSL and HFSH-2 sequence (LTPsub + LTPAP) in the BS-GCs (HFSH-1, 63.1 ± 9.6%, n = 8; HFSL-1 + HFSH-2, 108.5 ± 8.2%, n = 5, p < 0.05; Fig. 6F), whereas that in the RS-GCs was not significantly different from the LTPsub + LTPAP sum (HFSH-1, 39.6 ± 10.3%, n = 8; HFSL-1 + HFSH-2, 26.5 ± 12.9%, n = 5, p = 0.72). Importantly, LTP induced by HFSH-1 in BS-GCs was little affected by nimodipine (Nimo, 54.0 ± 10.8%, n = 6, p = 0.66; Fig. 6G). These results suggest that priming of BS-GCs by HFSL-1 is required for the contribution of L-VDCC to LTPAP induced by HFSH-2.
LTP induction at MPP-GC synapses is not affected by firing patternAbove results indicate that LTP at LPP-GC synapses can be induced by two distinct mechanisms: NMDAR-dependent subthreshold LTP and compound Hebbian LTP, and that the latter heavily depends on activation of L-VDCC resulting from postsynaptic AP bursts. We investigated whether MPP-GC synapses share the same LTP mechanisms with those of LPP-GC synapses. We recorded MPP-evoked baseline EPSPs using an electrode placed in the middle of the molecular layer in a 10 s interval for about 5 min before HFS was applied (Fig. 7A). The stimulation intensity was adjusted so that the peaks of EPSP summation evoked
留言 (0)