
記住我




Currently available claustrum marker genes, like Nurr1 and Nr2f2 (Additional file 1: Fig. S1A), are expressed, albeit sparsely, in the adjoining endopiriform nucleus, insula and gustatory-visceral cortex [3, 11,12,13], resulting in lack of claustrum specificity for these markers. Thus, there is a need for marker genes that separate claustrum cells from adjacent cortical cells. We reasoned that anatomical delineation of the claustrum could be achieved by leveraging the combinatorial molecular profiles of claustrum-enriched and cortical-enriched genes.
In a recent transcriptomic study, it was reported that a number of genes distinguish projection neurons in the claustrum from nearby cortical neurons [25]. We compared these findings with the Allen mouse brain in situ hybridization database [29] to identify candidate cortical-enriched genes that are devoid from the claustrum at early postnatal developmental stages and in adulthood (Additional file 1: Fig. S1B). We noticed that the transcription factor Tle4 satisfied both conditions better than other candidates: (1) single-cell transcriptomic molecular analysis shows that Tle4 is absent in the claustrum [25], and (2) Spatial expression of Tle4 appears to be highly enriched in deep layers of the insula as well as in deep layers of the sensory cortex, while avoiding the claustrum, both at P4 and P56 (Additional file 1: Fig. S1B). We identified an anti-Tle4 antibody that worked reliably for brain tissue derived from mice at different ages. Therefore, we set out to investigate whether Tle4 protein expression corroborates previous findings at the transcriptional level.
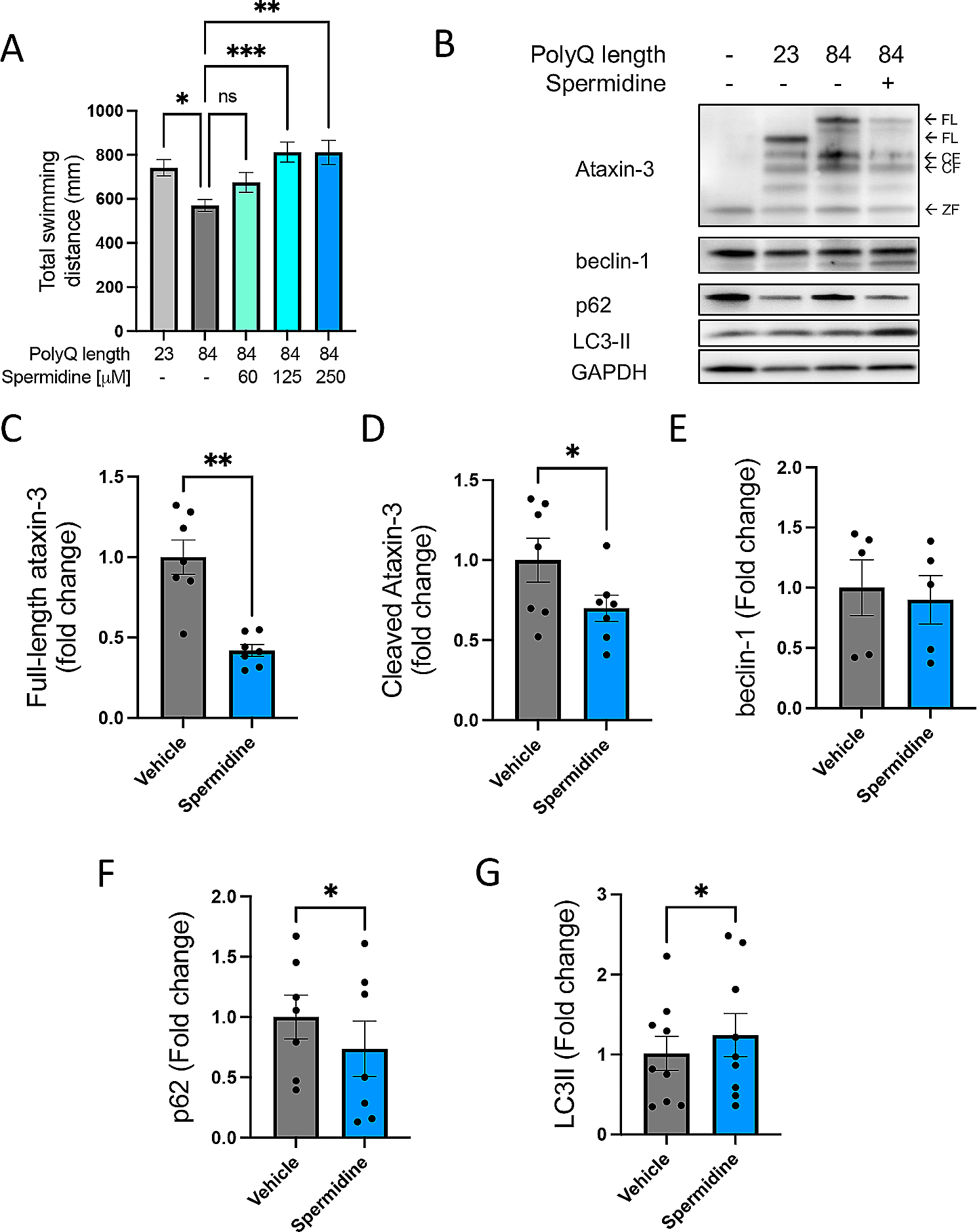
Topographical mapping of the claustrum has previously defined claustrum neurons projecting to the RSC as the center of the claustrum core, hereafter referred to as the central zone [15, 30]. Thus, for the first set of experiments, we used AAV retrograde fluorescent labelling of the RSC (Retro-RSC) as our reference to the claustrum. We performed unilateral injections of retro-AAV-CAG-GFP into the RSC of adult mice (> P70) and analyzed GFP expression in the claustrum 14 days post injection (Fig. 1A–C). Several studies found Nurr1 to be a relatively selective marker for the claustrum at different ages and across different species, including rodents and primates [10, 11, 13, 20]. We found that Nurr1 immunolabelling showed a dense population of cells that appeared to be spatially aligned with the GFP-labeled retro-RSC central zone (Fig. 1C, D). In contrast, Tle4 immunolabelling revealed that Tle4 was absent in the central zone, whereas outside of the central zone, Tle4 expression was noticeable in regions bordering the claustrum at all sides (Fig. 1E, F). This suggests that while Nurr1 is a claustrum-enriched marker, Tle4 is a potential claustrum-devoid marker. Next, we quantified colocalization of RSC-projecting cells with Nurr1 and with Tle4 in the anterior, middle and posterior subdivisions of the claustrum. While a large proportion of retro-RSC-projecting cells co-expressed Nurr1 (> 87%) (Fig. 1G, G’, I, J, L, M, O), very few cells co-expressed Tle4 (~ 1%) (Fig. 1H, H’, I, K, L, N, O). Of note, these results were consistent in all claustrum subdivisions along the anteroposterior axis, and thus verify absence of Tle4 expression in projection neurons throughout the central zone of the claustrum.
Fig. 1
The expression pattern of Nurr1 and Tle4 in the claustrum region distinguishes claustrum projection neurons from their cortical counterparts in layer 5 and 6. A Retrograde AAV-CAG-GFP was injected into the retrosplenial cortex (RSC), followed by tissue collection 14 days post injection. B An example tracer injection site in the retrosplenial cortex. C, E representative images of retrograde GFP labeling in the anterior claustrum (D, F), and representative images showing Nurr1 (D) and Tle4 (F) expression relative to GFP labeling. C–D are from the same slice, and E, F are from an adjacent slice of the same mouse. Dashed ellipses in (C–F) highlight GFP expression in the claustrum (CLA) without labeling the surrounding insula cortex (Ins) or striatum (str). G, H Representative images showing the degree of colocalization with Nurr1 (G) or Tle4 labelled cells (H). Single channel images are shown on the right of each panel. G’, H’ 3.5 × fold magnifications of areas in white boxes in (G, H), respectively, with single channel (left, middle) and merged (right) images. Orange arrowheads indicate examples of cell colocalization. I Venn diagrams showing the mean cell counts for Nurr+/GFP+ and Tle4+/GFP+ cells in the anterior CLA. Values throughout indicate mean ± standard deviation (n = 6 mice, 3 male and 3 female). J-L: The same as G-I but for the middle CLA. M–O The same as (G–I) for the posterior CLA. P Nurr1 and Tle4 colocalization in the CLA region was compared to layer 6b (Ctx L6b) of the primary motor cortex (MOp) and the somatosensory cortex (SSC). Q–S Representative images showing Nurr1 and Tle4 expression in the CLA region (Q), the MOp (R) and the SSC (S) (left, merged imaging channels; right, single channel images). Insets are 2.5 × fold magnifications of areas in white boxes. Red arrowheads indicate examples of cell colocalization. T–V Venn diagrams representing the mean number of cells expressing Nurr1 (magenta) and Tle4 (yellow), and colocalization between Nurr1 with Tle4 (n = 4 mice, 2 male and 2 female). The data in T are averaged across anterior, middle, and posterior CLA. Values in I, L, O, T, U, V represent mean ± standard deviation. Panels P–V: See Additional file 2: Table S1 for details on mouse sex
Outside of the claustrum, Nurr1 and Tle4 are found in deep layers of the sensory cortex, where Tle4 labels layers 5/6, and Nurr1 labels layer 6b [4, 31,32,33]. Therefore, we compared colocalization of Nurr1 with Tle4 in the claustrum to that in the cortex, namely in the motor and somatosensory cortices (Fig. 1P). Markedly, while only a small number of Nurr1-labeled claustrum cells co-expressed Tle4 (1.5%) (Fig. 1Q, T, Additional file 1: Fig. S2A, C, E), the vast majority of Nurr1-labeled cells co-expressed Tle4 (> 96%) in cortical layer 6b (Fig. 1R, S, U, V). As such, this stark difference in Tle4 molecular profile between Nurr1-expressing cells in the claustrum-insula complex versus the cortex can be used to discriminate claustrum projection neurons from nearby neuronal populations in cortical layer 6b.
In addition to Nurr1, Nr2f2 is another cortical marker for claustrum cells [12, 34]. Similar to Nurr1, there was an extensive overlap between Nr2f2-immunoreactive cells and GFP-labeled RSC-projecting cells in the central zone (Fig. 2A1, A2), and this overlap was maintained in claustrum subdivisions spanning the anteroposterior axis (> 81%) (Fig. 2B–G). However, GFP and Nurr1 co-expression was consistently higher than GFP and Nr2f2 co-expression across claustrum subdivisions (see Fig. 1). There was very low co-expression of Nr2f2 and Tle4 in the claustrum region (0.4%) (Fig. 2H, I, Additional file 1: Fig. S2B, D, F), thus confirming lack of colocalization between Tle4 and claustrum projection neurons in the central zone.
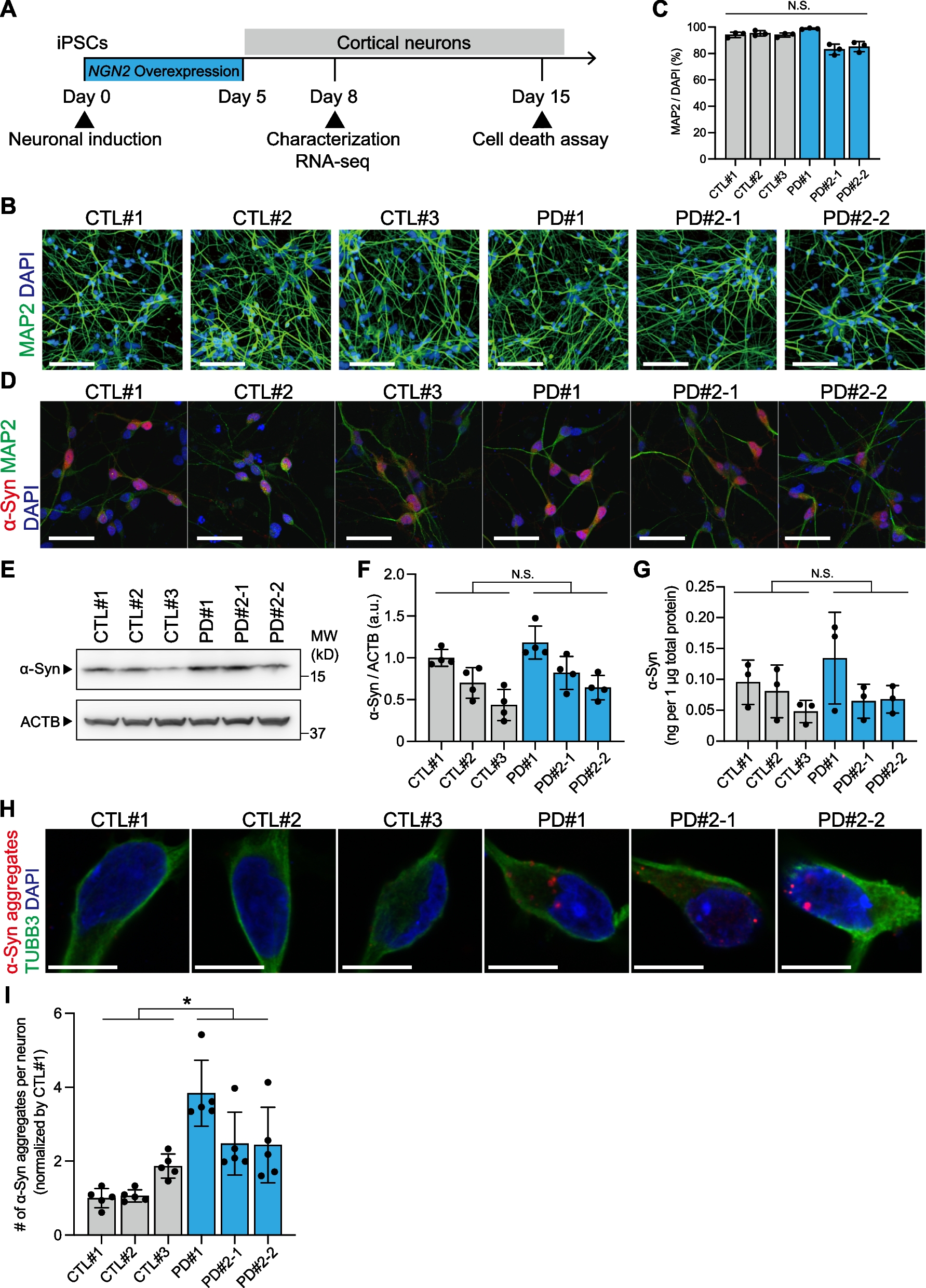
Fig. 2
Spatial pattern of Nr2f2 and Tle4 expression enables localization of claustrum projection neurons. A1–A2 Representative images of the anterior claustrum showing retrograde GFP labeling (A1) and Nr2f2 expression (A2) following AAV injection into the retrosplenial cortex (Retro-RSC). Dashed ellipses highlight GFP expression in the claustrum (CLA) without labeling the surrounding insula cortex (Ins) or striatum (str). B–D Representative images showing colocalization of GFP labeling with Nr2f2 in the anterior (B), middle (C) and posterior (D) CLA (left, merged imaging channels; right, single channel images). (B’) 2.3 × fold magnifications of area in the white boxes in (B) with single channel (top, middle) and merged (bottom) images. Orange arrowheads indicate examples of cell colocalization. E–G Venn diagrams representing the mean number of cells expressing Nr2f2 (red) and GFP (cyan), and colocalization of GFP with Nr2f2 in the anterior (E), middle (F), and posterior (G) CLA. Values indicate mean ± standard deviation from n = 6 mice (3 male and 3 female). H1, H2 Two representative images of the same coronal plane showing lack of colocalization between Nr2f2 and Tle4 in the CLA. H2 is a magnified image of the dashed box in (H1). For H2, merged imaging channels are on the left and single channel images are on the right. Inset (bottom-left) is 2.5 × fold magnifications of area in the white box. I Venn diagram representing the mean number of cells expressing Nr2f2 (red) and Tle4 (yellow), and the colocalization of Nr2f2 with Tle4, averaged across anterior, middle and posterior CLA (n = 6 mice, 3 male and 3 female). J1–J3 Example images of the CLA region showing polygons outlining the boundaries of GFP cells and neuropil (cyan, J1), Nr2f2 concentric expression (red, J2), and absence of Tle4 expression (grey, J3). K1, K2 Overlay of polygons from (J1–J3). Bottom-left insets show spatial registration of GFP outline with Nr2f2 outline (K1), and GFP outline with Tle4 outline (K2) (see Materials and Methods for details). Pink represents the topographic overlap of GFP/Nr2f2 expression, grey represents the region of low Tle4 expression. L Cumulative spatial overlap of GFP+/Nr2f2+ labeling (left), and the spatial location of GFP labeling within the region of relative Tle4 absence (GFP+/Tle4−, right), in the anterior (top), middle (middle) and posterior (bottom) CLA across 6 mice. Values in E, F, G, I represent mean ± standard deviation
In terms of spatial distribution, there was a dense group of cells in the claustrum central zone that exhibited strong Nr2f2 labelling when compared to the surrounding Nr2f2-immunoreactive cells, and this patch of Nr2f2-enriched cells seems to predominantly coincide with RSC-projecting claustrum cells (Fig. 2J1, J2, K1). On the other hand, the ring-shaped clustering of Tle4-immunoreactive cells around RSC-projecting cells appears to demarcate the perimeter of the claustrum (Fig. 2J1, J3, K2). The location of Nr2f2-enriched and Tle4-devoid domains relative to RSC-projecting cells was preserved across anterior, middle and posterior claustrum subdivisions, and was reproducible in different mice (Fig. 2L). Accordingly, these findings suggest a cell-type-specific spatial organization in the claustrum, with strongly labeled Nr2f2-positive cells occupying a relatively central domain along the dorsoventral axis of the claustrum, i.e. the central zone, and Tle4-expressing cells defining the perimeter of the claustrum.
Collectively, our data indicates that that Tle4 enrichment is exclusive to structures abutting the claustrum. Our data also shows that Nurr1 and Nr2f2 label the majority of claustrum cells in the central zone, albeit their labelling profile fails to delineate the boundaries of the claustrum. As such, combining Nurr1/Nr2f2 with Tle4 labelling highlights the contrast in spatial patterning between Nurr1/Nr2f2 and Tle4 within the claustrum region, thus providing an improved approximation of claustrum cell mapping.
Absence of Tle4 expression is a common feature for discrete subpopulations of claustrum cellsAre there neuronal populations devoid of Tle4 expression in the claustrum beside RSC-projecting neurons? To answer this question, we explored Tle4 colocalization with different subpopulations of claustrum projection neurons. It has been previously determined that claustrum neurons projecting to independent cortical regions are differentially distributed along the dorsoventral axis [15, 16]. To encompass the entire dorsoventral landscape of topographically positioned claustrum cells, we targeted different claustrocortical modules that are concentrated across the dorsoventral claustrum axis. To this end, we employed the same AAV approach as in the previous experiment to retrogradely trace claustrum cells projecting to the ACC (Fig. 3A, B), MOp (Fig. 3G, H) and LEC (Fig. 3M, N0, which comprise somewhat different claustrum populations in dorsal (Fig. 3C1, I1) and ventral zone (Fig. 3O1) relative to the central zone we determined above (see Fig. 2).
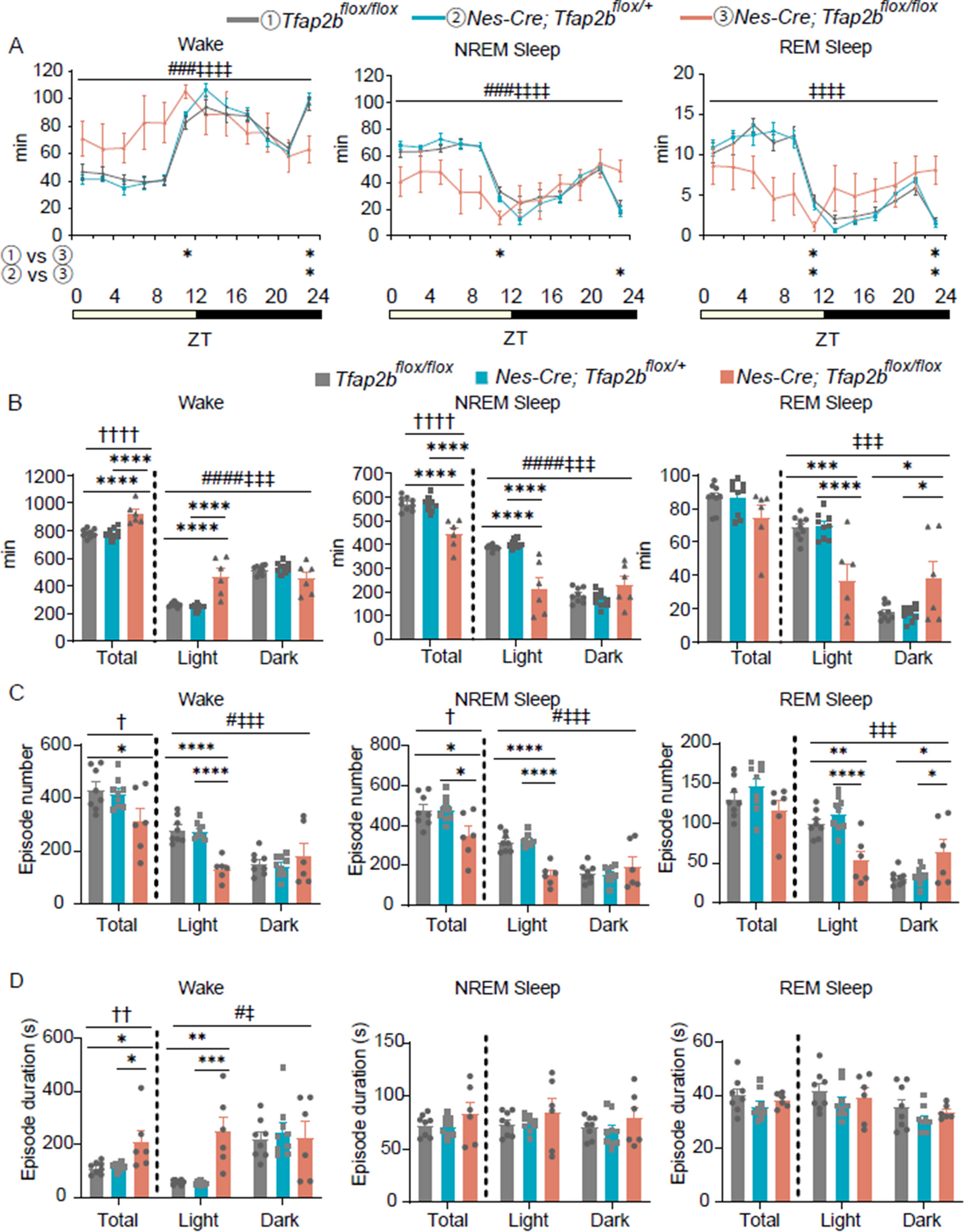
Fig. 3
Claustrum neurons projecting to different cortical regions exhibit colocalization with Nurr1 and Nr2f2, but not with Tle4. A Injection of retrograde AAV-CAG-GFP into the anterior cingulate cortex (ACC). Claustrum tissue was processed 14 days post injection. B Example of an AAV injection site in the ACC. C1–C4 Representative images from coronal sections of the anterior claustrum (CLA) relative to the insula (Ins) and the striatum (Str) showing retrograde GFP labeling following AAV injection the ACC (C1), along with the expression of Nurr1 (C2), Nr2f2 (C3), Tle4 (C4), and merged Tle4/GFP (C5). All panels are from the same slice, except Nurr1 which is from an adjacent slice. Dashed ellipses highlight the region of low Tle4 expression. D–F: Representative images (left) and Venn diagrams (right) showing colocalization of GFP labeling with Nurr1 (D), Nr2f2 (E) and Tle4 (F) in the anterior CLA. Panels E and F are the same slice. Venn diagrams in each panel show the mean number of cells expressing GFP with Nurr1 (D), Nr2f2 (E), and Tle4 (F) (n = 5 mice, all male). Values shown are the average cell counts across anterior, middle, and posterior planes of the CLA. Insets in left panels of D, E, F are 2.5 × fold magnifications of areas in white boxes. Orange arrowheads indicate examples of cell colocalization. G–L Same as A–F but for experiments with retrograde AAV-CAG-GFP injected into the primary motor cortex (MOp) (n = 4 mice, all male). M–R Same as A–F, but for retrograde AAV-CAG-GFP injected into the lateral entorhinal cortex (LEC) (n = 4 mice, all male). Values in right panels of D, E, F, J, K, L, P, Q, R represent mean ± standard deviation
We compared AAV labelling in each claustrocortical module with Nurr1, Nr2f2 and Tle4. For ACC-projecting cells, GFP expression was mainly confined between the central zone, demarcated by the enriched labelling of Nurr1 and Nr2f2 (Fig. 3C1–3), and the dorsal zone, delineated by the upper limit of Tle4-devoid region (Fig. 3C4, C5), thus corroborating our previous findings of ACC-projecting claustrum cells being preferentially located within central and dorsal zones [15]. On average, ACC-projecting cells were enriched in Nurr1 (77%), and to a lower extent in Nr2f2 (54%) (Fig. 3D, E), and they rarely expressed Tle4 (< 3%) (Fig. 3F).
Correlating the position of MOp-projecting cells with Nurr1 and Nr2f2 dense patches showed that MOp module corresponds to dorsal parts of the claustrum (Fig. 3I1–I3). Nevertheless, the Tle4 spatial distribution suggests that the module of MOp-projecting cells probably extends beyond the claustrum dorsal zone, raising the possibility that a subset of MOp-projecting cells is not part of the claustrum but rather belong to nearby dorsally located cortical structures (Fig. 3I4, I5). Quantification for the MOp-projecting module indicated that on average 61% of the cells co-expressed Nurr1 (Fig. J3), ~ 54% of the cells co-expressed Nr2f2 (Fig. 3K), and < 4% of the cells co-expressed Tle4 (Fig. 3L). These co-expression levels were somewhat similar to the ACC-projecting module.
LEC-projecting cells were clustered ventral to Nurr1 and Nr2f2 dense patches (Fig. 3O1–O3), and yet, these cells were contained within the lower limit of the Tle4-devoid area (Fig. 3O4, O5). This suggests that the LEC claustrocortical module is restricted to the ventral claustrum zone, which is in agreement with previously published data [15, 35]. While the colocalization of Nurr1 with LEC-projecting cells was comparable to ACC- and MOp-projecting cells (on average 72%, Fig. 3P), Nr2f2 colocalized with LEC-projecting cells at a considerably low level relative to ACC- and MOp-projecting cells (on average 29%, Fig. 3Q). Lastly, there was limited co-expression of LEC-projecting cells with Tle4 (on average 1.5%). Accordingly, Tle4 colocalization with all claustrocortical modules studied here was consistently low (< 5%), suggestive of a prominent association between lack of Tle4 expression and claustrum projection neurons across diverse claustrocortical modules. In each of the claustrocortical domains, i.e. ACC, MOp and LEC, there were similar proportions of colocalization between claustrum projection neurons with Nurr1, Nr2f2 and Tle4 (Additional file 1: Fig. S3).
We took a closer look at colocalization of Nurr1, Nr2f2 and Tle4 with RSC, ACC, MOp and LEC claustrocortical modules in the anterior, middle and posterior claustrum subdivisions. Overall, cells in the RSC module exhibited the most robust Nurr1 and Nr2f2 expression among all modules, and this was consistent across all anteroposterior subdivisions (Fig. 4A–F, Additional file 2: Table S1). In particular, differences between the RSC module and the ACC, MOp and LEC modules were significant in the anterior and middle claustrum with respect to Nurr1 (Fig. 4A, B) and in all subdivisions with respect to Nr2f2 (Fig. 4D–F). The only exception was Nurr1 expression in the posterior claustrum, where levels for the RSC module were significantly higher than in the MOp module, but not in ACC and LEC modules (Fig. 4C). Conversely, Nurr1 expression in the MOp module was consistently lower than in the ACC and LEC modules across all anteroposterior subdivisions (Fig. 4A–C). Yet, these differences were only significant in the anterior claustrum relative to ACC module (Fig. 4A), and the posterior claustrum relative to ACC and LEC modules (Fig. 4C). Also, Nr2f2 expression in the LEC module was lower than in the ACC and MOp modules (Fig. 4D–F, Additional file 2: Table S1), with differences being significant only between LEC and ACC modules in the middle and posterior claustrum (Fig. 4E, F). Thus, the expression of Nurr1 and Nr2f2 in claustrum cells varies between distinct claustrocortical modules, and also seems to be dependent on cell position along the anteroposterior axis. There were no significant differences in Tle4 levels between the different claustrocortical modules in any anteroposterior claustrum subdivision (Fig. 4G–I, Additional file 2: Table S1). These results provide evidence that distinct cell subsets in the claustrum are overwhelmingly devoid of Tle4 expression regardless of their dorsoventral or anteroposterior position.
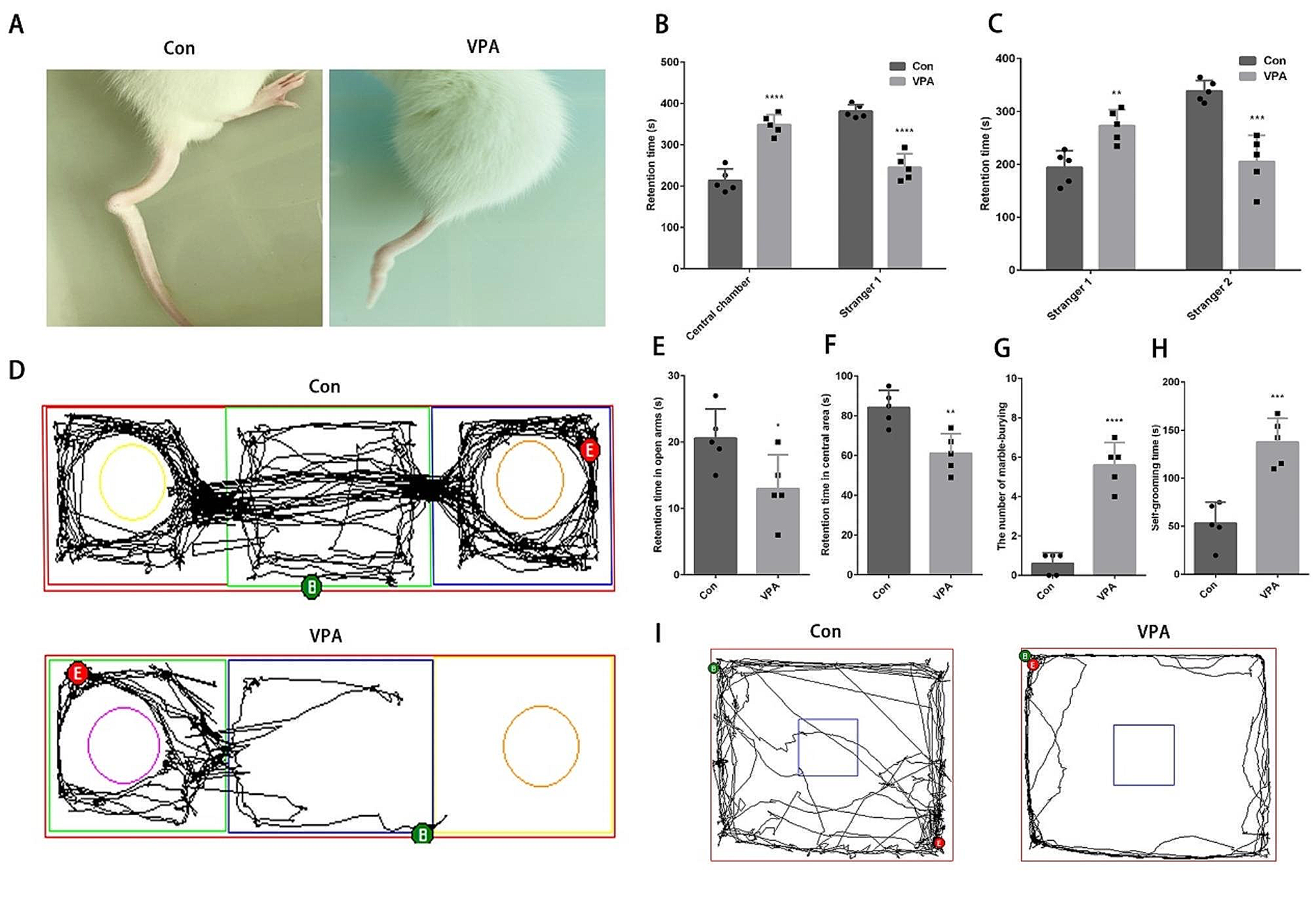
Fig. 4
Claustrum neurons projecting to the retrosplenial cortex express Nurr1 and Nr2f2 more extensively than other claustrocortical pathways. A–C Bar plots showing the mean percentage of GFP-expressing cells colocalized with Nurr1 (Nurr1+GFP+) in neurons projecting to the retrosplenial cortex (RSC), anterior cingulate cortex (ACC), primary motor cortex (MOp) and lateral entorhinal cortex (LEC) in the anterior (A), middle (B) and posterior (C) claustrum (CLA). D–F Same as A-C, but for GFP-expressing cells colocalized with Nr2f2 (Nr2f2+GFP+). G–I Same as A–C, but for GFP-expressing cells colocalized with Tle4 (Tle4+GFP+). Error bars represent mean ± SEM; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; one-way ANOVA followed by Bonferroni test (see Additional file 2: Table S2 for detailed statistical analysis). RSC (n = 6 mice, 3 male and 3 female), ACC (n = 5 mice, all male), MOp (n = 4 mice, all male), LEC (n = 4 mice, all male)
In summary, by selectively tracing separate claustrocortical modules, we were able to determine that not all projection neurons express the claustrum-enriched markers Nurr1 and Nr2f2. Interestingly, this heterogeneity of Nurr1/Nr2f2 expression in the claustrum appears to be spatially specific, where cells projecting from the central zone to the RSC comprise the least heterogeneous cell population. Our experimental approach demonstrates an overall minimal overlap between claustrum cells and Tle4-expressing cells. Thus, we have effectively established Tle4 as a bona fide marker for segregating the claustrum from adjoining cortical areas.
Claustrum projection neurons lack the expression of inhibitory marker genesIt was previously reported that Nurr1-expressing cells in the claustrum are excitatory neurons [25]. Because here we found that a sizeable portion of projection neurons in ACC, MOp and LEC claustrocortical modules did not express Nurr1 (30–40%), we sought to examine whether these cells have excitatory or inhibitory identity. For this purpose, we opted to focus on the ACC module since its spatial domain covers parts of the central and the dorsal zones (see Fig. 3).
PV and somatostatin (SST) inhibitory neuronal subtypes are known to make up 50–60% of claustrum interneurons [15, 36, 37]. Therefore, we performed co-immunolabelling of ACC-projecting cells in the claustrum with PV and SST markers. There was virtually no overlap between ACC-projecting cells and the expression of PV (~ 0.04%; Additional file 2: Fig. S4A–D) or of SST (~ 0.05%; Additional file 2: Fig. S4E–H). This suggests that projection neurons in the claustrum, identified using retrograde tracing, are predominantly excitatory neurons in line with many previous reports [24, 27, 38,39,40].
Tle4 molecular profile is suitable for identifying claustrum during early postnatal developmentRetrograde tracing is a widely used approach for mapping projection neurons during postnatal development. However, conducting tracing experiments in juvenile rodents to locate the claustrum is considerably demanding compared to their adult counterparts. Previous studies reported that locating the claustrum via PV plexus labelling of claustrum core or distribution of myelinated fibers surrounding the claustrum are less reliable before the beginning of the third postnatal week [20]. Based on Allen Institute mouse brain in situ hybridization database [29], Tle4 expression at early postnatal stages, i.e. P4, distinctly engulfs a small area in the ventrolateral cortex that is adjacent to the striatum, presumably the claustrum (Additional file 1: Fig. S1B). Accordingly, we determined if the Nurr1/Tle4 molecular profile we used to identify claustrum cells in adult mice is suitable for locating the claustrum during neonatal development. To test this, we injected retro-AAV-CAG-TdTomato into the ACC of mice at P0, P7 or P14 and analyzed virus expression after 7 days at P7, P14 or P21, respectively (Fig. 5A, B1–B3). It was logical to choose ACC as our injection site at such a young age since ACC retrograde tracing labels claustrum projection neurons at a higher density than RSC (our data and [20]). In addition, it was recently shown that CLA projections to the ACC develop earlier than RSC projections [20]. In order to compare the developmental results which used a seven-day virus expression time, an adult group was also injected at P49 and tissue was collected at P56 (Fig. 5A, B4). It is important to note that for P0, P7 and P14 mice, we adjusted injection volume to a smaller brain size to localize bolus delivery within the ACC and minimize non-specific viral spread to nearby regions (see Materials and Methods) (Fig. 5B1–B3).
Fig. 5
Claustrum cells in neonatal mice display the same Nurr1/Tle4 expression pattern as adult mice. A Injection of retrograde AAV-CAG-TdT into the anterior cingulate cortex (ACC) at different postnatal days, followed by tissue collection 7 days post injection. B1–B4 Examples of AAV injection sites in the ACC at P7 (B1), P14 (B2), P21 (B3) and P56 (B4) following injection at P1, P7, P14 and P49, respectively. C–F Representative images C, E showing the colocalization of TdTomato (TdT) labeling with Nurr1 (C) and Tle4 (E) in the anterior claustrum (CLA) at P7 (left, merged imaging channels; right, single channel images). Venn diagrams D, F representing the mean number of cells at P7 expressing Nurr1 and TdT (D), or Tle4 and TdT (F) (n = 5 mice). Values shown are the average cell counts from the anterior, middle and posterior coronal planes of the CLA. G–J Same as C–F, but for mice injected at P7 and perfused at P14 (n = 6 mice). K–N Same as C–F but for mice injected at P14 and perfused at P21 (n = 7 mice). O–R Same as C–F, but for mice injected at P49 and perfused at P56 (n = 5 mice). S Quantification of fluorescence intensity in the CLA region (cyan) in the medial–lateral axis (dashed line). T Measurement of fluorescence intensity (z-score) for TdT (cyan), Nurr1 (magenta) and Tle4 (black) labeling across the mediolateral axis of the CLA region at P7, P14, P21 and P56 (from left to right). Solid lines for each color represent the mean and light shaded areas of the same color represent standard deviation. U, V: Same as S–T, but for the dorsoventral axis. Insets in left panels of C, E, G, I, K, M, O, Q (bottom-left) are 2.5 × fold magnifications of areas in white boxes. Orange arrowheads indicate examples of cell colocalization. Values in D, F, H, J, L, N, P, R represent mean ± standard deviation (see Additional file 2: Table S3 for details on mouse sex)
At P7-P21, the number of fluorescently labeled ACC-projecting cells was lower than at P56 in the claustrum region (on average ~ 23 cells/slice at P7, ~ 27 cells/slice at P14, ~ 43 cells/slice at P21 and ~ 63 cells/slice at P56), yet co-localization of TdTomato-tagged cells with Nurr1 and Tle4 at all developmental ages was comparable to P56: TdTomato-Nurr1 > 80% at all ages; Tdtomato-Tle4 < 5% at all ages (Fig. 5C–R). Notably, the spatial distribution of most ACC-projecting cells at P7, P14 and P21 highly overlapped with Nurr1-enriched cell cluster in the claustrum region (Fig. 5C, G, K,), and at the same time ACC-projecting cells occupied the Tle4-devoid zone that is surrounded by Tle4-expressing cells (Fig. 5E, I, M). The same spatial patterns for both Nurr1 and Tle4 were replicated at P56 (Fig. 5O, Q), and were identical to our results in adult mice using retro-AAV-CAG-GFP (see Fig. 3). There was high colocalization of Nr2f2 and ACC-projecting cells between P7-P21 (65–88%), which was on par with the colocalization level detected in P56 mice (71%) (Additional file 1: Fig. S5). These data suggest that AAC-projecting cells in the developing claustrum display an analogous molecular profile for Nurr1, Nr2f2 and Tle4 to their counterparts in the adult claustrum.
We further measured the spatial density for marker expression within the claustrum region at P7, P14 and P21 relative to P56. As expected, the peak density of ACC-projecting, i.e. TdTomato-expressing, cells largely coincided with Nurr1-expressing cells across the dorsoventral and mediolateral axes at all analyzed ages (Fig. 5S–V). In contrast, the majority of Tle4-expressing cells were detected outside of the area with high TdTomato/Nurr1 fluorescence intensity from P7 and up to P56, with a bias towards being situated medial to as well as dorsal to the peak of TdTomato/Nurr1 fluorescence intensity (Fig. 5S–V). Thus, at all early postnatal stages of claustrum development, the spatial density of Tle4 expression displayed an inverse trend to TdTomato and Nurr1 expression. These results demonstrate that spatial distribution of Nurr1 and Tle4 is nearly identical in neonatal (P7-P21) and adult mice (P56), suggestive of a uniform Nurr1/Tle4 expression pattern throughout postnatal claustrum development.
We compared Nurr1/Tle4 expression between the claustrum and cortical layer 6b during early postnatal development (Additional file 1: Fig. S6). At P14, there was very low Nurr1/Tle4 colocalization in the claustrum (~ 0.8%) (Additional file 1: Fig. S6B, E), whereas layer 6b in the motor and somatosensory cortices displayed very high Nurr1/Tle4 colocalization (> 97%) (Additional file 1: Fig. S6C, D, F, G). These Nurr1/Tle4 colocalization results
留言 (0)