1. IntroductionIn December 2019, the outbreak of a novel coronavirus named severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) rapidly spread around the world, resulting in a global pandemic [
1]. Since then, international efforts to generate a suitable therapeutic have resulted in the development of multiple vaccination platforms and other antiviral pharmaceuticals. The gradual rise of variants has had a reduced impact on the efficacy of neutralising antibodies raised either by previous infection of SARS-CoV-2 or by vaccination [
2,
3]. The World Health Organisation (WHO) has categorised the troubling variants as variants of concern (VOC), whereas other variants that do not meet the same criteria fall under variants of interest (VOI) or variants under investigation (VUI). There has been a substantial amount of focus on variants and their characteristics, such as antibody evasion and replication rates, with many studies comparing variants and their ability to be neutralised [
4,
5,
6,
7,
8], as the pandemic continues to progress.SARS-CoV-2 belongs to the Coronaviridae family that includes SARS-CoV-1 [
9], middle eastern respiratory virus (MERS) [
10] and four human coronaviruses 229E, HKU-1, NL63, and OC43 [
11] (
Figure 1A). Whilst SARS-CoV-1 and MERS have had outbreaks that caused severe disease in humans [
12], the four other coronaviruses, commonly referred to as seasonal or human coronaviruses (HCoVs), typically cause mild disease similar to a common cold [
11,
13]. On rare occasions, however, the HCoVs may cause severe diseases [
14,
15,
16]. SARS-CoV-2, together with NL63, use angiotensin-converting enzyme 2 (ACE2) as their major cell entry receptor [
17,
18]. Despite HKU1 and OC43 being more closely related to SARS-CoV-2, they bind to sialic acids as a mode of entry [
19], whereas more distantly related 229E uses human aminopeptidase (hAPN) [
20]. (
Figure 1B).At the start of the pandemic, there was a debate as to the possibility that antibodies raised against the HCOVs had any role in protection against SARS-CoV-2 [
22,
23,
24,
25]. Since then, rising interest in HCoVs has led to an increase in understanding of the immune response they generate. Several publications that investigated the effect of HCoVs relied on the use of binding assays such as enzyme linked immunosorbent assays (ELISA) that measure antibody binding but did not elucidate their neutralising capabilities. Moreover, successful generation of various vaccine platforms have been used to protect individuals from infection and severe disease [
26], though their effectiveness is diminished as newer and more immune evasive variants arise [
27]. Here, we use lentiviral-based pseudotyped viruses of SARS-CoV-2, the VOCs/VOI, and HCoVs, to measure the strength of neutralising antibodies induced by two doses of either BNT162b2 (Pfizer), AZD1222 (Astrazeneca), or mRNA-1273 (Moderna) against SARS-CoV-2 and variants B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma), B.1.617.2 (Delta), B.1.525 (Eta), and B.1.1.529 (Omicron BA.1) (
Figure 2), and whether any of these vaccines are able to augment neutralising antibodies against HCoVs 229E, HKU1, or NL63. 4. Discussion
In this study, we were able to directly compare the antibody neutralisation titres induced by two m-RNA-based vaccines, BNT162b2 and mRNA-1273, and an adenoviral-based vaccine, AZD1222, against SARS-CoV-2, its emerged variants, and three seasonal HCoVs.
Our data on antibody neutralization against SARS-CoV-2 and its variants in vaccinated subjects, with or without a history of previous infection, agree with what is reported in the literature [
2,
4,
5,
7,
34,
35,
36,
37,
38]. We confirmed that vaccination with two doses of vaccines induced antibodies able to neutralize SARS-CoV-2 and VOCs, with BNT162b2 eliciting the highest neutralization titres, followed by mRNA-1273 and AZD1222. Despite their differences in neutralization titres, all three vaccines have been reported to have high efficacy at preventing severe COVID-19 [
39,
40,
41]. The Omicron BA.1 variant was the most evasive of all VOCs analysed in this study (
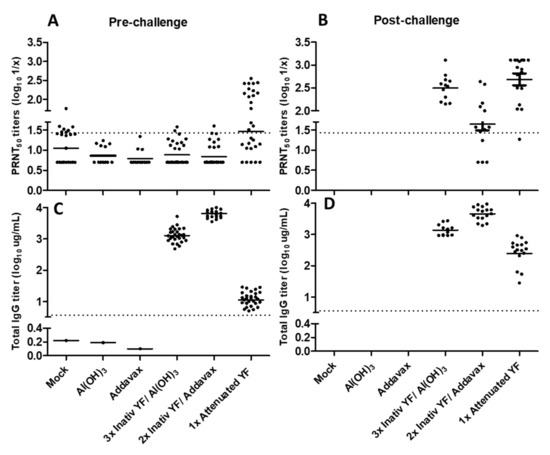
Figure 3). Indeed, the heavily mutated spike protein of BA.1 variant posed challenges to the effectiveness of the current vaccines to protect against COVID-19 and pointed out the need to monitor the protection conferred against this and the newly emerged SARS-CoV-2 variants, namely Omicron BA.4 and BA.5. Bivalent formulations of mRNA-based vaccines, containing both the mRNA of the spike of the ancestral SARS-CoV-2 and the one in common between the BA.4 and BA.5 lineages have been designed and authorized in order to counteract the evasion of the immune response elicited by the original vaccine design.HCoVs are globally distributed and believed to induce short-lasting protective antibodies [
42]. Therefore, there is a high likelihood of reinfection remaining elevated, especially during the winter periods [
13,
43,
44,
45] despite high seroprevalence [
43,
45,
46]. It is currently debated whether prior infection with seasonal HCoVs elicits cross-reactive antibodies against SARS-CoV-2, and more importantly, if this translates into protection against SARS-CoV-2. Cross-reactive antibodies [
47,
48,
49,
50,
51,
52,
53,
54] and T-cell responses [
55,
56,
57,
58,
59,
60,
61] were detected in pre-pandemic sera and healthy donors; however, similar experimental approaches have shown the opposite to be true by other investigators [
62]. In addition, in many of the aforementioned articles that revealed cross-reactive antibodies in pre-pandemic samples, the number of cross-reactive samples was a small portion of the total sera analysed, suggesting that cross-reactivity, whilst it exists, is low.The same question has been raised about antibodies elicited by COVID-19 vaccines, with studies showing cross-reactive antibodies to some but not all the seasonal HCoVs [
63,
64,
65]. SARS-CoV-2 spike protein vaccination was shown to induce cross-reactive antibodies to both Alpha- and Betacoronaviruses in macaques [
66]. It is important to deduce whether cross-reactive antibodies translate into protective, neutralising antibodies against SARS-CoV-2. Some reports suggested that whilst there is a small boost in antibodies towards HCoVs during SARS-CoV-2 infection, they are not associated with protection [
67]. Similarly, studies showed that prior infection with HCoVs did not protect against SARS-CoV-2 infection and disease [
68,
69].We did not find any boost of neutralizing antibody titres against HKU1 in our cohort of SARS-CoV-2-vaccinated subjects, irrespective of their SARS-CoV-2 pre-infectious status, with the exception of subjects administered with BNT162b2. This is in contrast with two reports that observed a boost in HKU-1 titres post vaccination against SARS-CoV-2 by BNT162b2 [
63,
64]. Hicks et al. showed that antibodies reacting to HCoV-OC43 and HCoV-HKU1 had minimal cross-reactivity with SARS-CoV-2, in accordance with the sequence homology of these proteins [
54]. Moreover, a previous SARS-CoV-2 infection did not boost the cross-neutralization against either HKU1 or the more phylogenetically related HCoV-229E (
Figure 4A). One report suggested that HKU1 may have another candidate receptor that has yet to be identified, due to the presence of a putative RBD, distant from the sialic acid binding regions [
70,
71]. It should also be noted that neutralising ability might not only be dependent on the pairwise similarity between amino acids in the protein, but also short insertions and deletions that can alter the protein’s structural conformation. For instance, the HKU-1-specific insertion at the C-terminal end of the RBD (
Figure 4C) might partly explain our neutralisation results.
Conversely, we found that the second dose administration in naïve subjects increased the protective antibody response against NL63 compared with that obtained in previously infected subjects receiving the same dose. This was probably due to the fact that additional exposures to the spike antigen did not have an effect on antibody neutralization against NL63.
The differences in antibody neutralization between the HCoVs may be due to the differences in the spikes used by the vaccination platforms. BNT162b27 encodes full-length spike with the K986P and V987P mutation sites to stabilize the pre-fusion conformation of the protein [
72]. The mRNA-1273 vaccine contains the coding sequence for a spike glycoprotein stabilized by the same proline substitutions used in the BNT162b2 vaccine, with a transmembrane anchor and an intact S1-S2 cleavage site. The pre-fusion conformation is stabilized by the consecutive proline substitutions, which are located in the S2 subunit at the top of the central helix [
73]. Conversely, a native-like spike is expressed by the AZD1222 vaccine. As our naïve subjects were administered with the AZD1222 vaccine, we can speculate that the native form of the spike protein triggered the development of higher neutralizing antibodies titres compared with that induced by the pre-fusion-stabilized protein.Conversely, the second immunogenic exposure to SARS-CoV-2 spike boosted the neutralizing response against NL63 or 229E (
Figure 4B), as has been previously reported [
67], depending on the vaccine platform, irrespective of the pre-infection status. Interestingly, another report observed the same cross-neutralizing activity, though this was irrespective of vaccine platform [
74]. We speculate that cross reactivity can arise due to similarity in epitopes in the receptor-binding motif (RBM) of NL63 to SARS-CoV-2, since both viruses share ACE-2 as their entry receptor [
75]. Similarly, an epitope overlapping the S2 fusion peptide in 229E has been reported to elicit cross-reactivity against SARS-CoV-2 [
48]. Song et al. described protective neutralizing antibodies targeting the S2 subdomain [
53]. Furthermore, a report during the original SARS-CoV-1 outbreak also found cross reactive antibodies against NL63 and 229E [
76], strengthening the hypothesis of shared epitopes between Alphacoronaviruses and Betacoronaviruses. The S protein of NL63 does not contain the furin-recognition site and is not cleaved during biogenesis [
77]. Similarly, the spike protein expressed by the mRNA1273 vaccine lacks the cleavage site; therefore, the conformation of the protein might be similar and might trigger neutralizing antibodies against shared epitopes and that are boosted after a second exposure to the same antigen.The antigenic nature of the spike protein expressed by the different vaccines, together with multiple conformations they can acquire, might affect the development of neutralizing antibodies with different affinities towards several epitopes in the spike protein. Since the AZD-1222 spike does not contain the two proline mutations to stabilise its spike into a trimeric pre fusion structure [
78,
79], the presence of a post fusion spike could potentially elicit a larger immune response towards epitopes in the S2 domain. This may explain why we did not observe any boost in neutralizing titres against 229E in either mRNA-based, pre-fusion-stabilized immunogen, vaccinated samples. Ultimately, despite observing a boost in titres, it is impossible for us to state whether this translates into protective titres since correlates of protection against SARS-CoV-2 have yet to be defined.
There are several limitations in our study to consider. Our data would have benefitted from larger numbers of samples in all vaccine platform types, and control samples of non-vaccinated individuals who either have been infected with SARS-CoV-2 or not. Furthermore, we did not analyse the baseline levels of cross-reactive neutralizing antibodies against seasonal coronaviruses in our cohort of vaccinated subjects.
A pan-coronavirus vaccine would elicit antibodies that recognise and neutralise a broad range of coronaviruses. This is challenging because of the genetic nature of these RNA viruses that frequently mutate and induce an immunity that wanes over time, increasing the likelihood of reinfection. Therefore, identifying the key epitopes located at the most conserved regions of the spike protein, especially at the S2 subunit, is relevant to potentially induce neutralizing antibodies with broader affinity to the cellular receptors that mediate viral entry. Several vaccine candidates have been formulated, and some are based on dual antigens including both spike and nucleocapsid (N) components [
80]. These formulations are at the pre-clinical stage as they might provide broader and more durable humoral and cellular immune responses against coronaviruses [
80].

留言 (0)