
記住我
Our current understanding of stress has evolved from early concepts pioneered by Hippocrates, who postulated that good health relied on maintaining a harmonious balance in physiological functions [1]. Building upon this idea, in the early 20th century, Walter B. Cannon, introduced the term “homeostasis“ to describe the coordinated physiological processes that maintain the organism’s steady states, expanding on Claude Bernard’s concept of the “Milieu intérieur“ from 1870. Although Cannon referenced “stress“ in his work, he never provided a formal definition [2].
Hans Selye extended Cannon’s notions of homeostasis and defined the word “stress” for the first time. He also scientifically measured the physiological changes it induces, although it is now recognized that his postulates do not apply to all species [3]. There is still a lack of consensus on defining stress, although Bruce McEwen’s perspective is widely embraced. “Stress may be defined as a real or interpreted threat to the physiological or psychological integrity of an individual that results in physiological and/or behavioral responses” [4]. His holistic approach also encompasses the positive and negative implications of stress responses [5], rendering it useful for application in preclinical and clinical research contexts.
The primary stress responses found in all vertebrates involve activating two key systems: the hypothalamic-pituitary-adrenal (HPA) axis and the sympatho-medullo-adrenal (SAM) axis. Activation of the HPA axis leads to increased levels of adrenocorticotropic hormone and glucocorticoids (such as cortisol in most mammals and corticosterone in rodents like rats and mice) in the bloodstream. The glucocorticoid receptors involved in regulating the HPA axis during stress are the glucocorticoid receptors (GR) and mineralocorticoid receptors (MR) [6]. Activation of the SAM axis results in elevated levels of catecholamines (like noradrenaline and adrenaline) in the bloodstream, and changes in the cardiovascular system and other bodily functions [7]. Fear responses activate these stress systems, leading to adaptative mechanisms such as hormonal and cellular events that help to confront or avoid the threatening stimulus [8].
Michael Davis’s definition of fear is broadly accepted, notably distinguishing it from anxiety, as it is crucial to differentiate between these closely related concepts. “Fear is prompted by imminent and real danger, and galvanizes active defensive responses. In contrast, anxiety is often elicited by less specific and less predictable threats, or by those that are physically or psychologically more distant” [9]. Joseph LeDoux advocates using the term fear only in people because it is intimately associated with the human consciousness. Therefore, he argues that fear should not be used in animals because we do not know how similar or different their consciousness is from human conscience. For experiments in animals, “threat conditioning” should be used instead of fear conditioning [10]. In literature, fear is often considered a more subjective state whereas threat-related behavior is a more descriptive term for observed behavior in model systems. In this review, fear represents both the subjective state and the objective threat-related behavior observed in animals and humans. One of the first reported fear conditioning experiments was performed in 1920, based on Ivan Pavlov’s conditioning principles [11]. In this experiment, an infant named Albert was exposed several times to a rat and a loud noise simultaneously. Following this initial exposure, the rat’s presentation alone caused Albert to cry (fear response) [12]. In 1928, Harrold Jones replicated and expanded upon these findings by incorporating psychogalvanic responses into analogous experiments. Similar to the skin conductance response (SCR), these responses pioneered objective measures of fear [13] leading to the foundation of the first fear conditioning laboratory. Some years later Estes and Skinner created the first procedure of Pavlovian fear-conditioning in rats and described the extinction of the conditioned responses [14]. Fear research conducted in the laboratory proves especially advantageous when studying memory mechanisms. This controlled environment allows significant control over the presentation of stimuli and the recording of psychophysiological, behavioral and neural responses.
Fear conditioning experiments typically involve an initial session (fear acquisition) that pairs an aversive stimulus (usually an electric shock) with a conditioned stimulus, like an acoustic tone or an image. An image is often used for humans, whereas, with animals, it is often an acoustic tone (cued-fear conditioning) or an experimental context (contextual fear conditioning). In a subsequent session (fear extinction), only the conditioned stimulus is presented without the electric shock, leading to the extinction of the fear response. In humans, the fear extinction test is frequently performed right after fear acquisition, but some studies test it 24 h later.
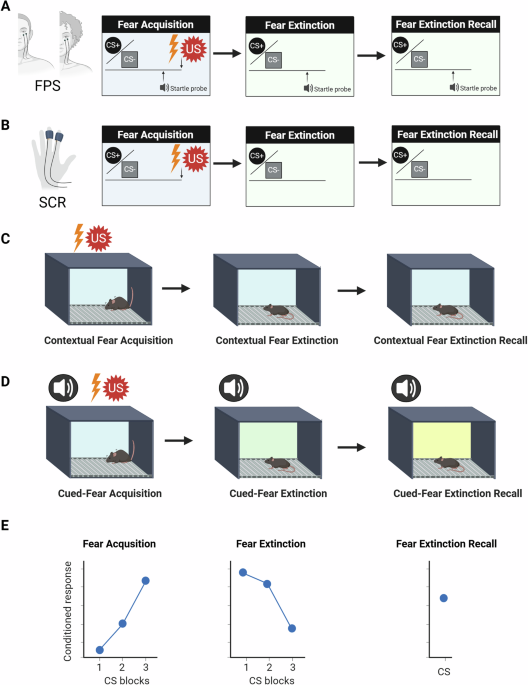
In animals, the fear extinction test is often performed 24 h after fear acquisition. Some studies on fear processing, especially on animals, also focus on recalling the initial conditioning (fear expression) [15]. The consolidation of the fear extinction memory is measured in the fear extinction recall session. See [15] for a methodological review with detailed explanations of different fear procedures in rodents and humans and a summary in Fig. 1. It should be noted that fear generalization is a process by which fear responses extend from a specific threatening stimulus to other similar but potentially harmless stimuli [15].
Fig. 1: A simplified summary of the fear learning paradigms most often used in humans and rodents.
A FPS fear-potentiated startle. US unconditioned stimulus, often an electric shock to the hand (annoying but not painful), an airblast directed towards the larynx or an aversive sound, especially used in children. CS conditioned stimulus. The CS+ is the stimulus paired with the US whereas the CS− is not paired with the US. The startle probe is a brief, sudden, loud noise that elicits an automatic response to close the eyes. The startle response is assessed by placing an electrode under the eye to measure action potentials with electromyography of the orbicularis oculi muscle. The pairing of the CS+ and the startle probe elicits an increase in the CS+ response. B SCR skin conductance response. US unconditioned stimulus, often an electric shock to the hand (annoying but not painful). CS conditioned stimulus. The CS+ is the stimulus paired with the US whereas the CS− is not paired with the US. In the non-dominant hand, this SCR measures emotional arousal to rewarding and aversive stimuli. FPS and SCR can be measured within the same experiment but often only FPS or SCR are measured. C US unconditioned stimulus, an electric footshock. A few mild electric footshocks are delivered in an experimental box. Animals are tested for fear extinction and extinction recall in the same experimental box keeping the context as similar as possible. D The cue is often an acoustic tone, an odor, or a light, which are paired with mild electric footshocks. US unconditioned stimulus, an electric footshock. The fear extinction and fear extinction recall are tested changing the conditions of the experimental box used for fear acquisition so that the rodent presents no contextual fear but cued-fear conditioning. For example, between the different fear phases, the color and lighting of the box should be changed. E The conditioned response is often the FPS or SCR in humans and freezing in rodents. CS blocks groups of CSs binned together.
An important advantage of fear conditioning experiments is that the neurocircuitry of fear is highly conserved across species, which allows the translation of results from rodent models to human research and the clinic [16]. Additionally, fear research can be combined with a wide array of techniques such as neuroimaging (e.g. functional magnetic resonance imaging, fMRI) in humans [15] and optogenetics, designer receptors exclusively activated by designer drugs (DREADDs) and freely moving calcium imaging recordings in rodents [17]. Despite using different methods and paradigms of fear conditioning in humans (fear-potentiated startle) and rodents (cued-fear conditioning) it has been shown that results can help to uncover shared molecular mechanisms of fear across species [18]. Morever, a recent study has found converging evidence of molecular mechanisms of fear in healthy mice and humans, using a combination of fMRI in humans and optogenetics, DREADDs, and freely moving calcium imaging recordings in mice [19]. Finally, to maximize the translatability of results, we must integrate the different methodologies, analyses, and conceptual frameworks used in humans and rodents to further understand fear memory processing [17].
Memory can be defined as the process by which knowledge of the world is encoded, stored, and later retrieved [20]. This conceptualization postulates that memory involves two stages: short-term memory, which lasts minutes or hours, and long-term memory, which can last days, weeks, or years. The main difference is that long-term memory requires the induction of genes and proteins, while short-term memory does not. Importantly, stress is a key modulator of fear memory mechanisms with implications in the clinic of fear-based disorders such as post-traumatic stress disorder (PTSD) [21] and PTSD-like animal models [17]. The prevalence of PTSD is around 5,6% in trauma exposed individuals but highly variable depending on many factors such as the type of trauma and peritrauma experienced [22, 23]. Examples of trauma can be a car accident, sexual abuse, military combat and personal assault among others. Women and individuals identifying as trans-masculine present higher vulnerability to PTSD compared to other demographics [23, 24], although the causes are not well understood yet.
Fear acquisition, extinction, and extinction recall in humans with PTSD or rodents exposed to traumatic stress compared to healthy individuals can be quite different [17]. The most consistent result of traumatic stress exposure on fear memories has been generally described as impairment of both fear extinction and extinction recall. Mixed results are found regarding traumatic stress exposure and fear acquisition suggesting either similar or enhanced levels compared to healthy individuals. These results may depend on many factors, including the nature of the traumatic stress and sex differences, with females being more vulnerable to enhanced fear acquisition [25,26,27,28,29,30].
Previous literature provides an understanding of the fear mechanisms of healthy rodents and humans (e.g. [31, 32]). Other reviews have extensively covered data concerning neural activity in various brain regions related to PTSD, stress and fear memory (e.g. [33]).
In this review, the main focus is on the molecular mechanisms of fear memory in patients with PTSD, not healthy individuals, and its relevant animal models. These appropriate models are rodents exposed to acute traumatic stress (PTSD-like models) with subsequent exposure to fear conditioning and extinction. See Fig. 2. Chronic traumatic stress in animals is beyond the scope of this review, as it induces significant components of depression-like behavior in addition to impaired fear extinction [33, 34]. Consequently, chronic traumatic stress may serve as a valuable model for studying PTSD comorbid with depression.
Fig. 2: Fear memories in healthy individuals and individuals exposed to traumatic stress.
Studies in rodents and humans agree on fear memory after exposure to traumatic stress, leading to impaired fear extinction and fear extinction recall. Figure inspired by Maren and Holmes [33].
As most studies test fear conditioning and extinction in healthy animals and rarely incorporate these methods with traumatic stress exposure, the available literature is limited. Moreover, when the sex of the animals is not specified in this review, it must be assumed to be male subjects, as they have been the most used historically in research. Finally, more efforts should be placed into testing both male and female subjects, as data in neuroscience can present striking sex differences [35]. CS blocks groups of CSs binned together.
Sex differences in stress interacting with fear memoriesAn important characteristic of PTSD is the inability to extinguish fear in safe environments, leading to impaired fear extinction when using FPS [36] or SCR [37]. Additionally, intrusive distressing thoughts in PTSD are associated with impaired fear extinction in FPS [38]. Patients with PTSD also present impaired extinction recall [39], which has been studied in both men and women, mainly with SCR.
When comparing sexes, women with PTSD show similar or enhanced levels of fear acquisition [40, 41] and men show deficits in fear extinction recall [41] measured with SCR. Both men and women with PTSD and healthy individuals generally present a higher response to the CS+ compared to the CS− [41].
Many acute and chronic stress procedures in rodents induce impaired fear extinction, such as restraint, forced swimming, maternal separation, predator exposure, social defeat and isolation, elevated platform exposure, single-prolonged stress (SPS) and stress immobilization on a board (IMO) [33]. For SPS, rodents undergo a 2-h restraint followed directly by a 20-min forced swim. After a brief recovery period, they are exposed to diethyl ether until they reach a state of anesthesia and become unresponsive [33]. In IMO, rodents are placed in a prone position with the limbs attached to metal mounts on boards [42]. IMO is different from the restraint used in SPS and IMO elicits a greater HPA axis response than other types of less severe restraint methods [42]. E.g. restraint in a tube. Moreover, IMO elicits a more prolonged HPA activation than intense and repeated electrical footshocks in rats [43].
Typically, all these different acute and chronic stress procedures are applied some hours or days before the fear procedures, allowing time for the incubation of PTSD-like symptomatology (e.g. [44]). However, most of these methods have only been combined with fear conditioning procedures in male rodents, showing overall impaired fear extinction. Several studies have also investigated female rodents, and there are findings regarding the stress-induced impaired fear extinction phenotype that do not consistently align with those observed in male rodents highlighting significant sex differences. E.g. SPS [33, 45,46,47,48,49]. However, both males and females previously exposed to IMO present equivalent fear acquisition levels and impaired cued-fear extinction [29, 30]. Moreover, IMO has been used to uncover fear molecular mechanisms of stress-related processes in rodents that may be relevant to PTSD in studies involving data in mice and humans [18, 29].
Finally, when studying stress interacting with fear memories, the differences seen between sexes can be attributed to their underlying mechanisms, which involve sex hormones, genetics, epigenetics, and other factors that are beyond the scope of this review.
Hormones influencing stress and fear memory mechanismsSome studies directly measure hormonal levels in blood or saliva, and others indirectly by monitoring the menstrual (women) or estrous cycle (female rodents). Rebecca Shansky´s work highlights the importance of considering hormonal levels, especially sex hormones, in preclinical and clinical settings to improve the quality of neuroscience research [35]. Here, I will focus on the most studied hormones important in traumatic stress and subsequent fear memory processing: estradiol and cortisol/corticosterone.
We have recently reviewed the role of estradiol and other relevant sex hormones, such as progesterone and testosterone, in fear memories in healthy individuals [31]. Most of the estradiol is produced by the granulosa cells of the ovaries. This process involves the aromatization of androstenedione (produced in the theca follicular cells) to estrone, followed by the conversion of estrone to estradiol by the enzyme 17β-hydroxysteroid dehydrogenase [50].
Seminal work by Mohammed Milad established that high estradiol levels enhance fear extinction retention in healthy women and female rodents (See the review [31]). This effect warrants further research in women with PTSD especially when considering their menstrual cycle or endogenous estradiol levels [51, 52]. For example, women diagnosed with PTSD presenting low and high estradiol levels in their blood showed similar levels of fear acquisition with FPS [51]. Moreover, women with PTSD and low estradiol levels show impaired fear extinction [53]. Also, women in the midluteal phase of the menstrual cycle (high progesterone and estradiol levels) diagnosed with PTSD show impaired fear extinction retention using SCR [51]. Ethinylestradiol is a synthetic estrogen widely used in oral hormonal contraceptives (HCs) and it appears to enhance fear extinction in women with PTSD. Women exposed to trauma and exhibiting various PTSD symptoms and using HCs exhibited increased acquisition of fear conditioning and enhanced fear extinction compared to women not using HCs and men [54]. Estradiol has been proposed as an adjunct to prolonged exposure therapy involving fear memories in women with PTSD [55]. Finally, the picture of fear memories and estradiol in patients with PTSD is complex, and further studies are needed to link all the different data.
Studies in rodents exposed to traumatic stress before fear procedures evaluating the role of estradiol are minimal. Recent data suggests that the menstrual or estrous cycle at the moment of traumatic stress (IMO) is not associated with impaired fear extinction in mice and PTSD symptoms in women exposed to sexual abuse [29]. These suggest that endogenous sex hormones or the menstrual/estrous cycle during traumatic stress may not be as important as previously suggested for the development of PTSD [56]. However, exogenous sex hormones could be beneficial after traumatic stress to normalize fear memories. For example, female ovariectomized rats showed that systemic injections of β-estradiol after exposure to the SPS reduced contextual freezing suggesting potential therapeutic effects [57]. Interestingly, estradiol plays a complex role in modulating cortisol and corticosterone levels in rodents and humans, with effects dependent on the species, sex, and context of exposure [58,59,60].
In general, cortisol and corticosterone enhance fear memory consolidation in healthy individuals but impair delayed memory retrieval [61, 62]. However, opposite-sex effects of these glucocorticoids on fear memories in healthy humans and rodents have been described and should be further explored. In people, hydrocortisone impaired the contextualization of fear expression, leading to increased fear generalization in FPS data among women, whereas the opposite pattern was observed in men [63]. In mice, systemic corticosterone enhanced cued-fear memory consolidation in males whereas it impaired it in females [64]. It needs to be noted that the HPA axis in rodents and humans although it has many similitudes also presents important differences among species and sexes [65] that must be considered when translating findings across species.
Similar to other hormones like estradiol, cortisol exhibits different effects on fear memories between healthy individuals and patients with PTSD [66]. Interestingly, patients with PTSD often show low baseline cortisol levels [67], and there is extensive research suggesting that giving glucocorticoids in the aftermath of traumatic stress could prevent the development of PTSD [68]. However, the potential therapeutic effect of administering glucocorticoids in patients with PTSD to normalize fear memories is still a complex issue that depends on many variables, including the timing and the dose of the glucocorticoid. For example, a single glucocorticoid administration to patients with PTSD appears to reduce the recall of fear memories but enhances their extinction measured with SCR [66, 69]. Moreover, it has been shown that using the potent GR agonist dexamethasone reduces exaggerated fear responses to the CS+ during FPS in patients with PTSD [70]. Considering altered baseline levels of cortisol and a likely non-linear relationship between corticosteroid levels and memory function, it may be that stress-induced cortisol brings healthy individuals and patients with PTSD to different places on an inverted U. However, this does not necessarily mean that glucocorticoids are doing different things across groups of individuals, but the effects of fluctuating corticosteroid levels must be interpreted in the context of these baselines. It is also possible that altered glucocorticoid levels in individuals with PTSD may not only alter memory strength, but generalization.
Results in animal models align with findings in humans, but unfortunately, these studies have only been performed on male subjects. For example, IMO leads to increased corticosterone levels during cued-fear conditioning and extinction and impairment of fear extinction [71]. Interestingly, inhibiting corticosterone synthesis with metyrapone during fear memory consolidation disrupted exti
留言 (0)