
記住我
Rheumatoid arthritis (RA) is an autoimmune and inflammatory disease characterized by the involvement of multiple cells and cytokines in its pathogenesis. In the RA synovium, various cellular phenotypes have been identified, including myeloid, lymphoid, low inflammatory, and fibroid, each with a distinct gene expression signature (1). Notably, higher levels of T cells and B cells have been observed in the synovium of anti-CCP positive RA patients compared to anti-CCP negative patients (2). Cutting-edge laboratory technologies such as single-cell transcriptomics and mass cytometry analysis have further revealed several synovial phenotypes in RA (3).
Multiple advanced therapies targeting a specific pathway in the pathogenesis of RA have been developed. However, there are no reliable markers to guide the optimal use of these medications. Consequently, many patients with moderate to severe RA undergo “trial and error” process to identify the most suitable treatment option for their condition. Therefore, there is an unmet need for effective biomarkers that help assess the efficacy and predict the response of RA medications.
CXCR5- PD-1hi peripheral helper cells (Tph) have been described in synovium of seropositive RA patients (4), in contrast to CXCR5+ T follicular helper cells (Tfh), which function predominately in the lymph nodes. One of the primary functions of both Tfh and Tph cells is to promote differentiation of B cells into antibody-secreting plasma cells.
Abatacept, a fusion protein consisting of the extracellular domain of human CTLA-4 and a fragment of the Fc domain IgG1, works by blocking an interaction between CD80/CD86 on antigen presenting cells and CD28 on T cells. Abatacept has been reported to be more effective in seropositive RA patients with high baseline anti-CCP levels (5) and HLA-DRB1 risk alleles (6).
It has been shown to suppress Tfh cell maturation and proliferation in murine models of RA (7). Additionally, abatacept treatment reduced circulating Tfh cells and lowered the expression of the activation marker ICOS on T cells in patients with primary Sjogren’s syndrome (8) and multiple sclerosis (9). In RA patients, abatacept has been shown to reduce serum ACPA and RF levels and lower the frequencies of post-switch memory B cells (CD19+CD27+IgD-) (10). These results suggest that the inhibition of activated Tfh and Tph cells, leading to attenuated T cell-dependent B cell activation, may underlie the efficacy of abatacept. CD19+CD11c+ (IgD-CD27-) double negative 2 B cells emerged as autoreactive B cells that are activated in extrafollicular pathways (11) and are increased in patients with rheumatoid arthritis and other rheumatic diseases (12). In addition to serving a precursors of antibody secreting cells (ASCs), DN2 CD11c+ B cells have been also proposed to present antigens to T cells (13) and interact with Tfh/Tph cells (14). Still little is known about the potential effects of abatacept on suppressing DN2 B cells in connection with its direct effects on reducing Tfh and Tph cells.
Soluble markers shed from activated T cells and B cells have been reported to be increased in various inflammatory conditions including RA (15–17). Additionally, cytokines and chemokines associated with Tfh and Tph cells such as IL-21 and CXCL13 were elevated in RA (18, 19). Their levels may reflect activity of specific cell population and could be useful markers to evaluate RA inflammation. It is unknown whether these markers improve in response to abatacept.
The intent of this study is to investigate the pathways leading to B cell activation and autoantibody production and identify novel biomarkers that could be helpful in predicting and assessing the response to abatacept in RA patients who have an inadequate response to methotrexate and other conventional disease modifying anti-rheumatic drugs (cDMARDs).
Materials and methodsStudy populationA total of 25 adult patients with RA who were recruited from the rheumatology clinics at the University of Washington, Seattle, WA, USA were enrolled between May 2019 and December 2022 (Table 1). All the participants fulfilled the 1987 ACR criteria or 2010 ACR/EULAR criteria for RA (20) and had clinical disease activity index (CDAI) ≥16 at screening corresponding to moderate to severe disease activity. The subjects had an inadequate response to conventional DMARDs including methotrexate, leflunomide, hydroxychloroquine, and/or sulfasalazine within the past year and were biologic-naïve except one patient who took etanercept 8 months prior to screening and stopped due to adverse events.

Table 1. Baseline characteristics.
All the subjects received abatacept 125mg subcutaneous injection weekly for 24 weeks and continued conventional DMARDs which they were on at screening visit. The subjects had 4 visits (baseline, week 6, week 14, and week 24) and peripheral blood samples were obtained at each visit. Responders to abatacept were defined as subjects who achieved ACR50 response at week 24. The research protocol was approved by the Institutional Review Board of the University of Washington, and all patients provided written informed consent for participation in the study.
Flow cytometryPeripheral blood mononuclear cells (PBMCs) were isolated with Ficoll-Hypaque (GE Healthcare Life Sciences, Marlborough, MA) as instructed. After isolation, PBMCs were stored frozen with Cell Freezing and Preservation Media (STEMCELL Technologies, Vancouver, Canada) in liquid nitrogen until further use. Vials of cryopreserved PBMC were thawed at 37°C in a water bath before immediately being diluted drop-wise into warm RPMI 1640 containing 10% FCS (Fisher Scientific, Waltham, MA). Thawed PBMCs were stained with combinations of fluorescently labeled antibodies (Supplementary Table 1) to identify distinct T cell and B cell subsets (Supplementary Figures 1, 2). All longitudinal samples from the same subject were run on the same day. Samples from two to six subjects were run on the same day with the same setting template. Live cells were identified using the Zombie Fixable Viability Sampler Kit (BioLegend, San Diego, CA). Ten-parameter flow cytometry was performed and analysed for changes in different T cell and B cell subsets at baseline, week 14, and week 24 using a 4-laser CytoFLEX flow cytometer (Beckman Coulter, Brea, CA). An average of 300,000 live lymphocytes were collected, and samples with <50 events for evaluated populations at baseline were excluded from the longitudinal analysis. All data were analysed using FlowJo software (Tree Star, Ashland, OR).
ELISAsFreshly isolated serum samples were aliquoted and stored in a −80°C freezer until use. Commercial enzyme-linked immunosorbent assay (ELISA) kits were used for serum measurements of soluble CD23 according to the manufacturer’s instructions (R&D Systems, Minneapolis, MN). Anti-CCP ELISA Assay Kits (Eagle Biosciences, Amherst, NH) were used to determine the IgG levels of anti-CCP antibodies and Rheumatoid Factor IgM ELISA kits (Abcam, Cambridge, UK) were applied to measure serum IgM RF as instructed by the manufacturers. Anti-CCP and IgM RF were also measured via multiplex and turbimetry in the clinical laboratory (Table 1) at University of Washington Medical Center. The upper limit of anti-CCP was 300 U/mL in the clinical lab.
Luminex immunoassaysThe following biomarkers in serum, including soluble CD25, soluble CD27, soluble CD40L, soluble PD-1, CXCL10, CXCL13, and IL-21, were measured with ProcartaPlex by Luminex Immunoassays (Thermo Fisher Scientific, Waltham, MA) at the Histology and Imaging Core, University of Washington (Seattle, WA) (https://uwhistologyandimaging.org/histopathology-work-order/).
HLA-DRB1 genotypeHuman leukocyte antigen (HLA)-DRB1 shared epitope (SE) genotypes were determined using targeted next-generation sequencing on frozen PBMCs (LabCorp, Burlington, NC). Patients were considered SE-positive if they had ≥ 1 of the following HLA-DRB1 alleles (6): *01:01, *01:02, *01:05, *04:01, *04:04, *04:05, *04:08, *04:09, *04:10, *04:13, *04:16; *04:19, *04:21, *04:35, *04:66, *10:01, *14:02, *14:06, *14:09, *14:13, *14:17, *14:19, *14:20, and *14:21. Patients were classified as SE2 (2 SE allele), SE1 (1 SE allele), or SE0 (no SE alleles).
Statistical analysisGraphPad Prism 8.0 (GraphPad, San Diego, CA) and SPSS software were used for statistical analyses. For sample sets with a non-Gaussian distribution of values, the Mann-Whitney test and Spearman’s correlation test were used as applicable. Categorical variables were compared using Chi-squared test or Fisher’s exact test. Changes before and after treatment were compared using the Wilcoxon matched-pairs signed rank test. The associations between baseline variables and response to abatacept were estimated by Lasso regression analyses. P values less than 0.05 were considered significant.
ResultsClinical efficacy of abataceptTwenty-three patients completed the study. One patient dropped out because of concerns for COVID infection and one patient dropped out for a reason that was not related to the study. The baseline characteristics of the patients are summarized in Table 1. Responders to abatacept were defined as subjects who achieved ACR50 response at week 24. After 24 weeks of treatment, 10 patients (43%) had ACR50 response. ACR20 response was achieved in 14 patients (61%) and ACR70 response was achieved in 3 patients (13%). Seven patients (30%) achieved DAR28-CRP remission (<2.6). There were no significant differences between responders and nonresponders though the frequency of females tended to be higher and anti-CCP and RF positivity tended to be lower among nonresponders. Consistent with prior work (5) where abatacept was more effective in seropositive RA patients with high anti-CCP levels, nine out of ten patients with ACR50 response had high baseline anti-CCP levels (>300 U/ml) and were double-positive for both anti-CCP and IgM RF (Table 1). In this study, 10 out of 23 subjects (43%) experienced adverse events. There were 3 upper respiratory infection, 3 headaches, 3 gastrointestinal symptoms (1 abdominal pain, 1 diarrhea, 1 intestinal infection) and 1 urinary tract infection. There were no serious adverse events and no subject discontinued abatacept because of the adverse events.
Comparison of baseline markers between responders and nonresponders to abataceptWe compared the baseline levels of T cell and B cell related soluble and cellular markers between responders and nonresponders to assess whether these markers are associated with response to abatacept. In patients who achieved ACR 50 response at 24 weeks, baseline levels of anti-CCP, RF, CXCL13, sCD23 and frequencies of CD11c+IgD-CD27- B cells were elevated as compared to patients not achieving ACR50 response (Table 2). The increase in sPD-1 levels among responders did not reach statistical significance (p=0.06). Baseline IL-21 levels were elevated only in patients who had ACR20 response (p=0.03, data not shown). On the other hand, there was a tendency that sCD25 were lower at baseline (p=0.09) in patients who had ACR50 response.
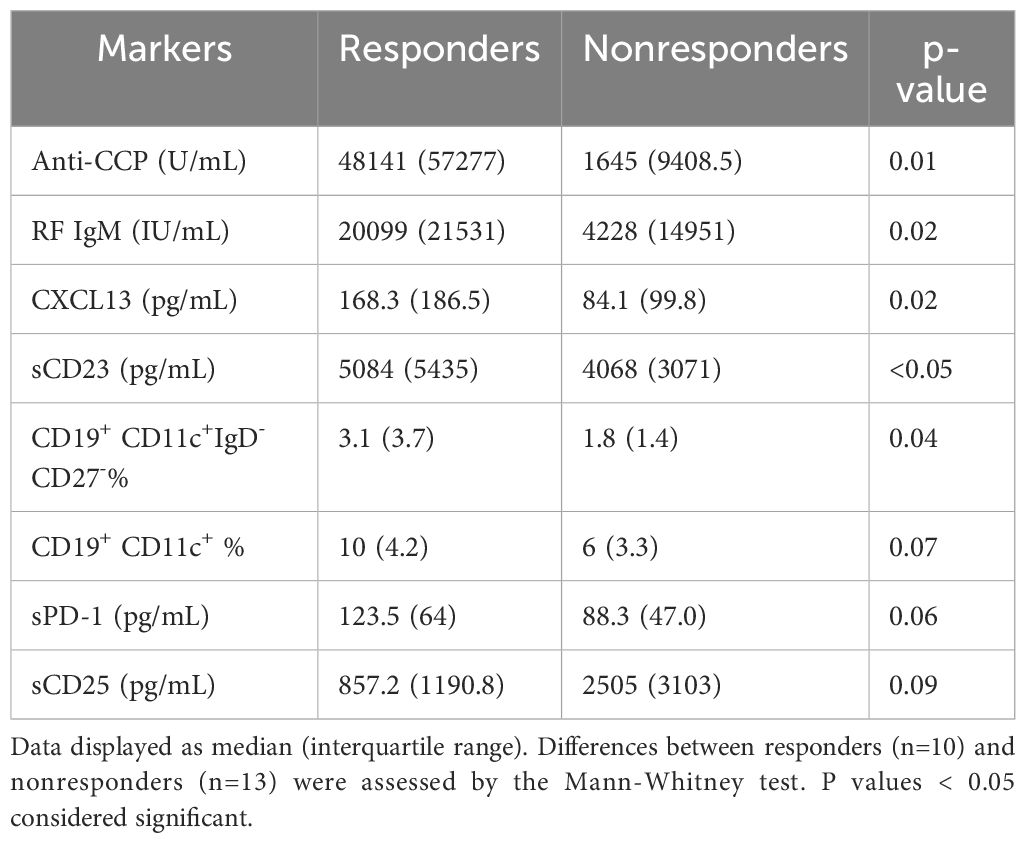
Table 2. Comparisons of baseline markers between responders and nonresponders to abatacept.
Immunologic markers associated with autoantibodiesOur data showed a strong correlation between anti-CCP and IgM RF levels (r=0.76, p<0.0001). To further identify the pathways related to autoantibodies in seropositive RA, we investigated potential links between multiple immunologic markers and anti-CCP and IgM RF (Table 3). We found that Tfh and Tph cell related markers (CXCL13, IL-21, sPD-1) were associated with anti-CCP or IgM RF. Of note, most of these markers including sPD-1 correlated more strongly with IgM RF compared to anti-CCP. Among T cell subsets, frequencies of CD4+CXCR5+PD-1hi Tfh cells and CD4+CXCR5-PD-1hi Tph cells were associated with both anti-CCP and Ig RF. Among B cell subsets, frequencies of CD19+IgD-CD27- B cell and CD19+CD11c+IgD-CD27- B cell were associated with anti-CCP but not with IgM RF. In Lasso regression, we found levels of anti-CCP and frequencies of CD19+CD11c+IgD-CD27- B cells independently associated with the response to abatacept. Notably, the frequencies of CD19+CD11c+IgD-CD27- B cells appeared to be more strongly associated with the response to abatacept as compared to anti-CCP, as indicated by their higher coefficient (Supplementary Table 2).
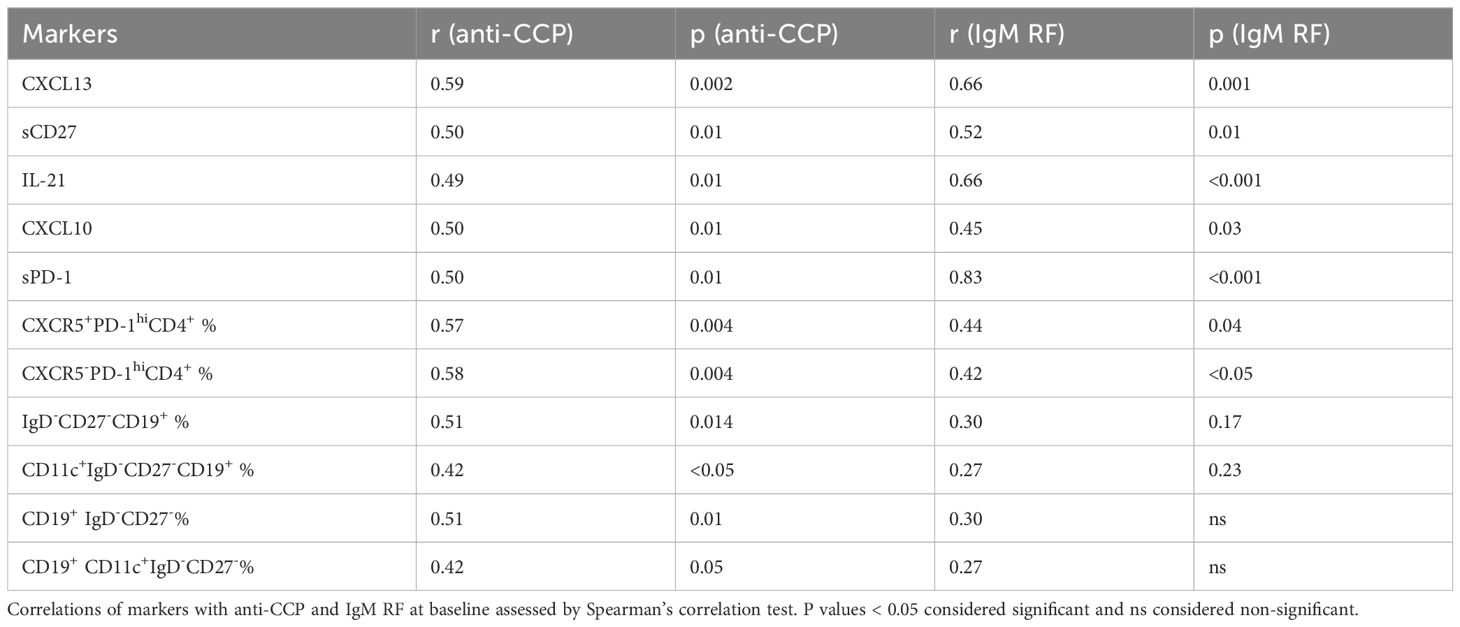
Table 3. Markers associated with anti-CCP and IgM RF in all RA patients (n=23).
Shared epitope related to response to abataceptBesides immunologic markers, RA patients were also genotyped for HLA-DR SE alleles and classified as SE2 (≥ 2 SE allele, n=8), SE1 (1 SE allele, n=11), and SE0 (no SE alleles, n=4) (Figure 1A). The presence of two alleles of shared epitope (SE) was associated with ACR50 response (p=0.04, Figure 1B) and was not significantly associated with ACR20 response (p=0.09, data not shown). Patients with 2 alleles of shared epitope had higher levels of anti-CCP (p=0.02, Figure 1C) and IgM RF (p=0.04, Figure 1D) when compared to patients with 1 or no SE allele.
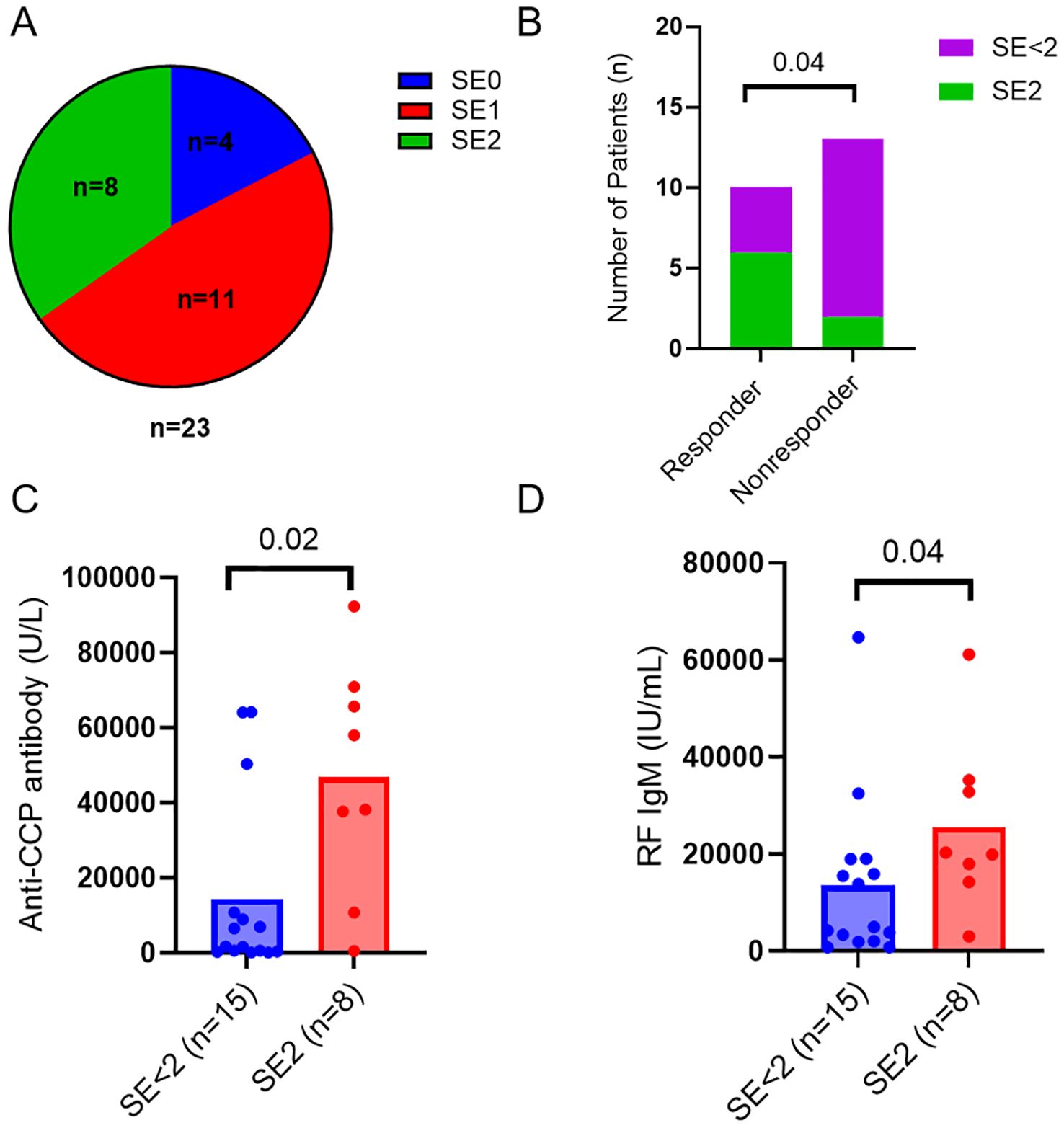
Figure 1. HLA-DRB1 shared epitope related to response to abatacept. (A) Pie chart showing numbers of RA patients with different SE alleles in the cohort. (B) Numbers of RA patients with two SE alleles (SE2) or less (SE<2) in responders and nonresponders. Differences between groups were determined by Fisher’s exact test. Baseline serum levels of anti-CCP antibodies (C) and RF IgM (D) in the RA patients with 2 alleles of shared epitope (SE2) and the control patients with 1 or no SE allele (SE<2). Differences between groups were determined by the Mann-Whitney U test. P values < 0.05 were considered significant.
Longitudinal changes in T cell and B cell related soluble and cellular markersWe assessed the changes of frequencies of different immune cell phenotypes and levels of soluble markers to determine the effect of abatacept treatment on these markers. There was a significant decrease in frequencies of circulating CD4+CXCR5+PD-1hi Tfh cells (Figure 2A) and CD4+CXCR5-PD-1hi Tph cells (Figure 2B) at week 14 and week 24. There was also a significant reduction in other T cell subsets, including CD4+CD25+CD127- regulatory T (Treg) cells, CD4+CD45RA-CCR7+ central memory T cells, and CD4+CD45RA-CCR7- effector memory T cells (Supplementary Table 3). Furthermore, abatacept decreased ICOS expression on both Tfh cells and Tph cells at week 14 (p<0.0001 and p<0.0001, respectively) and week 24 (p<0.0001 and p<0.0001, respectively) (Supplementary Figure 3). Abatacept also decreased frequencies of circulating CD19+IgD-CD27+ class-switched memory (SM) B cells, CD19+CD27+CD38hi plasmablasts/plasma (PL) B cells, CD19+CD11c+ B cells and CD19+CD11c+IgD-CD27- B cells at week 14 and week 24 (Figures 2C–F; Supplementary Table 4) as well as CD11c expression on B cells at week 14 and 24 (p=0.01 and p=0.006 respectively) (Supplementary Figure 4). Serum levels of sCD23, CXCL13 and RF IgM were significantly decreased at week 14 and week 24 with the treatment of abatacept when compared to baseline, whiles sCD27, CD40L and IL-21 levels were only decreased at week 14, sCD25, and sPD-1 were only reduced at week 24 (Table 4). Changes of anti-CCP levels did not reach statistical significance (Table 4).
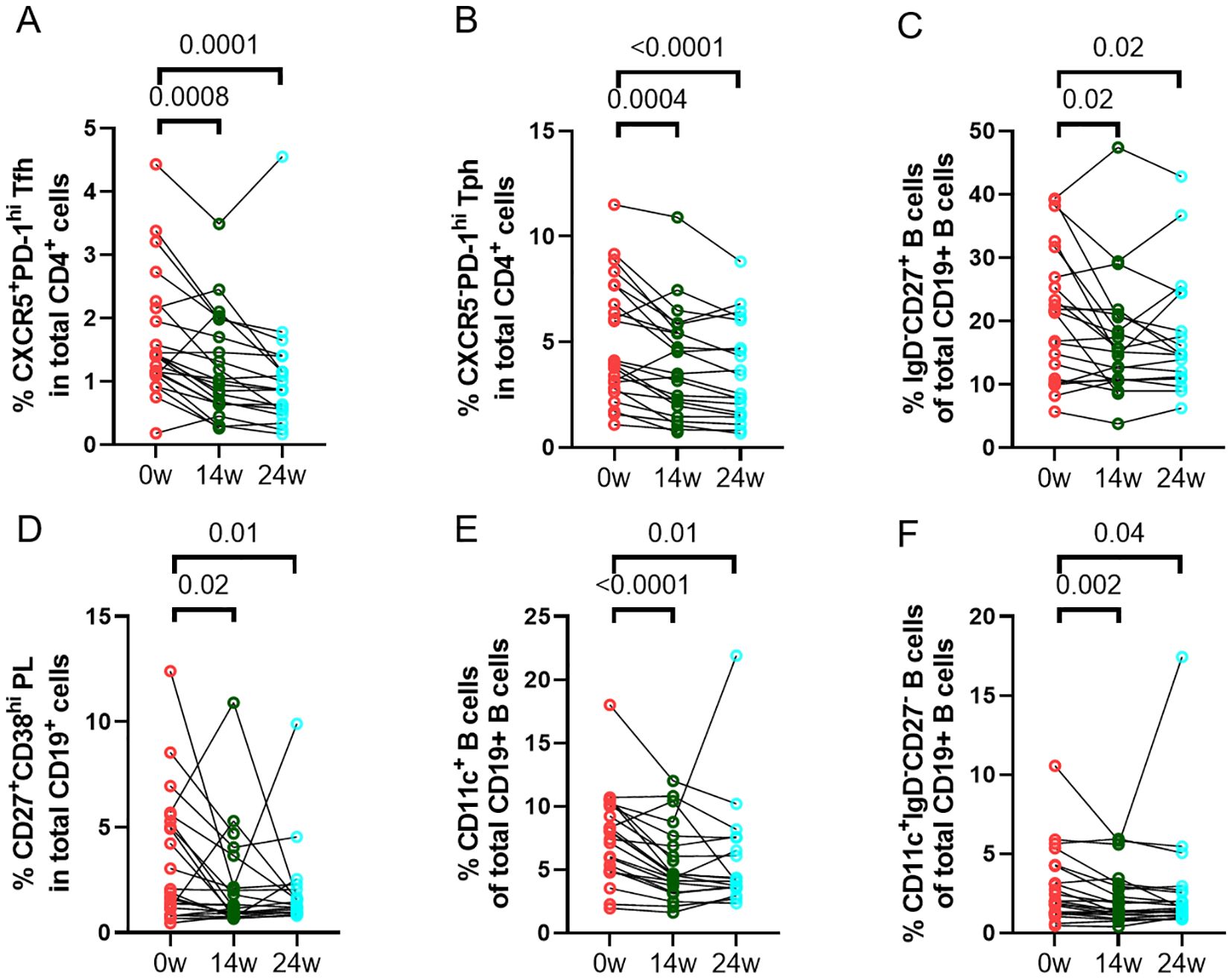
Figure 2. Changes in percentages of immune cell phenotypes. Frequencies of CD4+CXCR5+PD-1hi Tfh cells (A), CD4+CXCR5-PD-1hi Tph cells (B), CD19+IgD-CD27+ class-switched memory (SM) B cells (C), CD19+CD27+CD38hi plasmablasts/plasma (PL) B cells(D), CD19+CD11c+ B cells (E), and CD19+CD11c+IgD-CD27- B cells (F) in all RA patients (n=23) were documented before and after treatment with abatacept. Differences between baseline and post-treatment (week 14 and week 24) were determined by the Wilcoxon matched-pairs signed rank test. P values < 0.05 were considered significant.
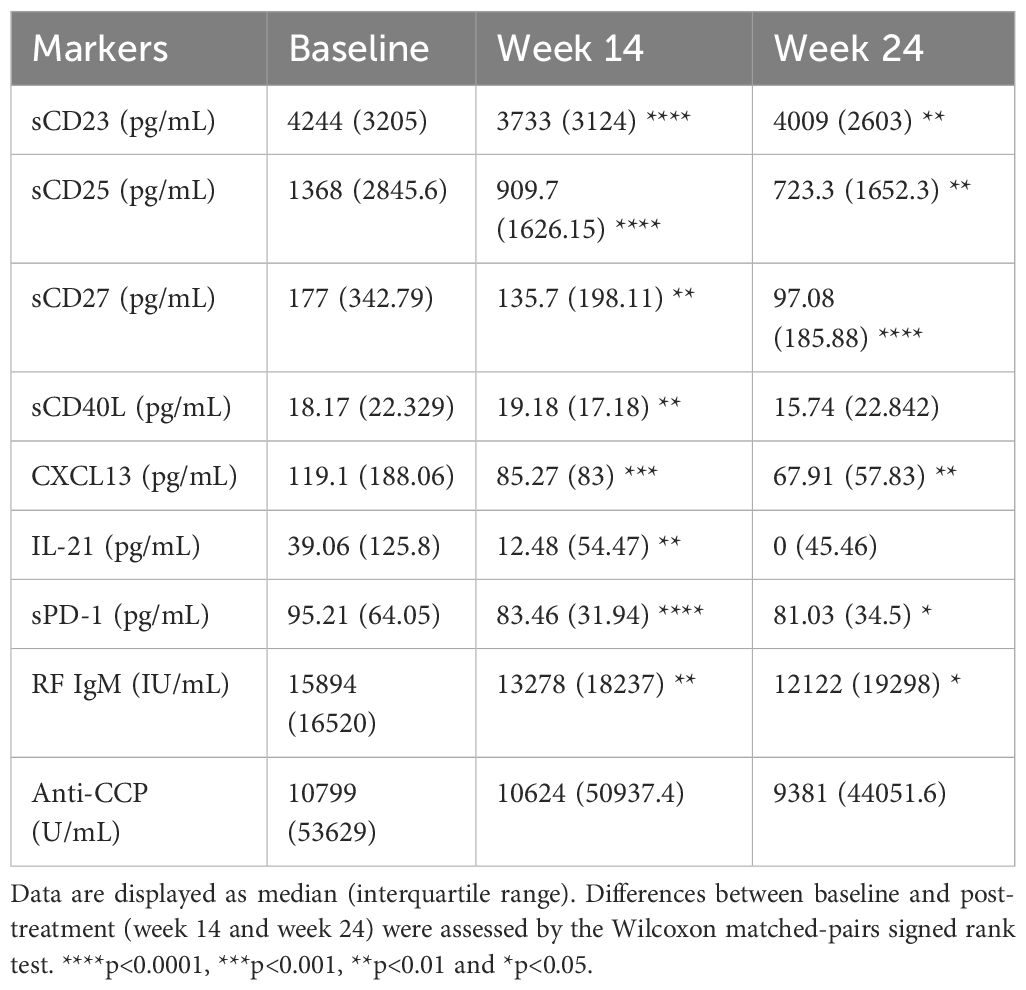
Table 4. Levels of soluble markers before and after treatment with abatacept in all RA patients (n=23).
CXCL13 and IgM RF as pharmacodynamic biomarkers for abatacept treatmentWe compared the longitudinal changes in T and B cell related markers including IgM RF and anti-CCP levels between responders and nonresponders. There were differences in longitudinal alterations in CXCL 13 (Figure 3A) and IgM RF (Figure 3B) levels between responders and nonresponders. CXCL13 and IgM RF levels decreased only in responders who achieved either ACR50 response (p=0.004 and p=0.02, respectively, at week 24) or ACR20 (p=0.03 and p=0.006, respectively, at week 24, data not shown). Frequencies of CD19+CD11c+IgD-CD27- B cells only decreased in responders at week 14 (p=0.02) but not at week 24 (Figure 3C). On the other hand, sCD23 (Figure 3D) were reduced significantly in both responders and nonresponders, while anti-CCP levels (Figure 3E) did not decrease significantly in either responders or nonresponders.

Figure 3. Serum CXCL13, RF IgM but not anti-CCP levels decreased in responders with treatment of abatacept. Data are displayed as a mean of serum levels of CXCL13 (A), RF IgM (B), CD11c+CD27-IgD- B cells (C), sCD23 (D), and anti-CCP antibodies (E) before and after treatment of abatacept in responders (n=10) and non-responders (n=13), error bars display the 95% confidence. Differences between baseline and post-treatment (week 6, week 14, and week 24) were determined by the Wilcoxon matched-pairs signed rank test. P values < 0.05 were considered significant. **p<0.01, *p<0.05, ***p<0.001 and ns considered non-significant.
DiscussionIn this study, we have reported several immune markers which are associated with the response to abatacept in RA patients. Though majority of patients had some response with abatacept, we chose the ACR50 response at week 24 as the responder criteria to identify the markers that are associated with a major improvement in disease activity from abatacept therapy. Consistent with previous studies (5, 6), abatacept was more effective in seropositive rheumatoid arthritis (RA) patients with positive anti-CCP and RF.
There have been new approaches to try to predict the response to biologic agents based on synovial biopsy phenotype in RA (1, 21). However synovial biopsy is not available nor standardized in most of rheumatology clinics currently and peripheral blood markers would be preferred by many clinicians, not least due to blood sampling being less invasive. In this study, T and B lymphocyte related markers including CXCL13, IL-21, sPD-1 and sCD23 were significantly decreased with the treatment of abatacept. CXCL13, IL-21 and PD-1 are expressed by T follicular helper (Tfh) cells and T peripheral helper (Tph) cells infiltrated in RA synovium (3) which play crucial roles in stimulating B cell activation and autoantibody production.
Similar to our findings, levels of CXCL13 and sPD-1 levels have been shown to correlate with RF titers in RA patients (19, 22). CXCL13 is involved in recruiting CXCR5+ B cells to synovium in RA and was associated with disease activity (23), ultrasonographic synovitis (15) and long-term radiographic progression (24). CXCL13 is mainly secreted by follicular dendritic cells and Tfh cells (25) and abatacept decreases its levels by suppressing activation of these two cells through interrupting the interaction between CD80/86 and CD28. Additionally, high CXCL13 levels were observed in RA patients with the lymphoid phenotype in synovium (1). These patients responded better to abatacept as shown in our study. Thus, though validation is required, CXCL13, reflecting T- and B-lymphocyte-driven synovial inflammation, may be a novel blood-based marker of treatment response to abatacept.
Our data showed that baseline frequencies of CD19+CD11c+IgD-CD27- B cells were higher in responders to abatacept. These cells are commonly referred to as double-negative (DN) 2 B cells (11) and have been identified in contexts of infection (26) and autoimmune diseases (27), where they are often termed atypical memory B cells or age-associated B cells (ABCs), as characterized by the expression of the T-box transcription factor (T-bet) (28, 29). It’s been revealed that CD11chiCD27lowT-bet+ B cells are primed to differentiate into plasma cells and produce autoantibodies targeting RNA-associated antigens such as Sm and SmD in SLE (30). In RA, DN2 B cells are major precursors to synovial antibody-secreting cells (31). To our knowledge, this is the first study to show an association between CD19+CD11c+IgD-CD27- B cells and response to abatacept in RA. Additionally, we found that CD11c levels on B cells decreased with abatacept treatment. Tfh cells have been reported as a major source of IL-21 in various inflammatory and autoimmune diseases (32–34). In alignment with these findings, our data also showed that abatacept treatment significantly reduced serum IL-21 levels in RA patients. It’s been reported that IL-21 can drive the expansion of CD11chiT-bet+ B cells (DN2) and promote their differentiation into Ig-producing autoreactive plasma cells (30) and that IL-21 and interferon-γ (IFNγ)-coexpressing PD-1high CXCR5-HLA-DR+ Tph cells skewed B cell differentiation toward a CD21low CD11c+ phenotype in vitro (35). Based on these findings, we hypothesized that CD28 blockade by abatacept disrupts the development and maintenance of the Tfh and Tph cells, thereby suppressing DN2 B cell expansion through decreased IL-21 production.
The percentage of B cells expressing CD23, the low-affinity receptor for the Fc portion of IgE, and serum levels of soluble CD23 (sCD23) were increased in patients with RA (16). Soluble CD23 is cleaved from naïve B cells when these cells are matured to memory B cells and the levels of sCD23 decreased with B cell depletion therapy, rituximab, in RA patients (36). We have shown abatacept was more effective in RA patients with a higher sCD23 levels which reflected B cell activation and differentiation. Our findings are thus consistent with the known mechanism of action of abatacept, inhibiting Tfh and Tph cell activation that leads to suppression of B cell differentiation.
On the other hand, soluble CD25 (sCD25) levels were numerically higher in nonresponders. Soluble CD25 is a high-affinity soluble IL-2 receptor and can form a complex with IL-2. This cytokine-receptor complex can potentially activate various cells including CD4 and CD8 effector T cells and NK cells through “trans presentation” of IL-2 to the cells (17). We found that sCD25 levels were lower in RA patients with 2 alleles of shared epitope (data not shown). Based on those observations, sCD25 may be associated with a distinct pathway of immune cell activation for seronegative RA that is not responsive to B cell modulation by abatacept.
Furthermore, RA patients with two shared epitope (SE) alleles had a higher probability to achieve ACR50 response from abatacept therapy and there seemed a close relationship between presence of shared epitopes and anti-CCP levels. Though a previous study showed abatacept resulted in higher efficacy responses versus adalimumab after 24 weeks in SE-positive patients (6), this is the first study to demonstrate two SE alleles being associated with treatment response to abatacept. This genetic marker will add additional information in evaluating RA if used along with anti-CCP levels as the latter may change with treatment.
Lastly, we found CXCL13 and IgM RF levels decreased with treatment only in responders to abatacept whereas anti-CCP levels did not decrease in either responders or nonresponders. Abatacept would inhibit production of autoantibodies including RF by suppressing Tfh and Tph cells. Given that we measure IgM isotype of RF which are secreted by newly activated B cells, IgM RF levels would be more preferably decreased by abatacept compared to IgG autoantibodies. Our results were consistent with the report that rheumatoid factor levels correlated with disease activity better than anti-CCP in RA patients (37). We plan to conduct an additional analysis of RA clinical trial data to determine if these markers could be used to monitor the response to abatacept.
The limitations of this study include subject number and study population. First and foremost, our study had a small number of subjects recruited from a single center and our results should be validated in a larger RA cohort. It would be useful to assess these markers in a specific RA population including early-onset RA or anti-CCP high RA patients. Additionally, our study population was primarily patients who had an inadequate response to DMARDs and biologic-naïve. It would be interesting to evaluate if these markers are also associated with the response to abatacept in patients who had inadequate response to biologic agents including TNF inhibitors. The exact function of DN2 B cells in RA, their interaction with T cells, and their BCR repertoire, are beyond the scope of this current study. However, there is a significant interest in this B cell subset in RA which should be further evaluated in the future studies.
In conclusion, we have shown the markers of B cell activation including frequencies of CD19+CD11c+IgD-CD27- B cells in RA were associated with favorable response to abatacept. Further, RA patients with two copies of shared epitope had a higher probability to achieve ACR 50 response. Though multiple markers related to immune cell activation decreased with abatacept therapy, CXCL13 and IgM RF decreased only in responders and could be potentially used as pharmacodynamic markers for abatacept in RA treatment.
Data availability statementAll relevant data is contained within the article: The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statementThe studies involving humans were approved by Institutional Review Board of the University of Washington. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributionsTW: Data curation, Formal analysis, Writing – original draft, Writing – review & editing. NG: Conceptualization, Supervision, Writing – review & editing. CL: Data curation, Formal analysis, Writing – review & editing. NW: Formal analysis, Writing – review & editing. BH: Writing – original draft, Writing – review & editing, Conceptualization, Funding acquisition, Supervision.
FundingThe author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Bristol-Myers Squibb. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Conflict of interestBH received a research grant and consulting fees from Bristol-Myers Squibb. CL received research grants from Eli Lilly, Gilead Sciences, Pfizer, Boehringer Ingelheim, Redd Pharmaceuticals and Amytrx, and is on the scientific advisory board for Redd Pharmaceuticals and Citryll.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statementThe author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1504454/full#supplementary-material
References1. Dennis G, Holweg CTJ, Kummerfeld SK, Choy DF, Setiadi AF, Hackney JA, et al. Synovial phenotypes in rheumatoid arthritis correlate with response to biologic therapeutics. Arthritis Res Ther. (2014) 16:R90. doi: 10.1186/ar4555
PubMed Abstract | Crossref Full Text | Google Scholar
2. Orr C, Najm A, Biniecka M, McGarry T, Ng CT, Young F, et al. Synovial immunophenotype and anti-citrullinated peptide antibodies in rheumatoid arthritis patients: relationship to treatment response and radiologic prognosis. Arthritis Rheumatol. (2017) 69:2114–23. doi: 10.1002/art.v69.11
PubMed Abstract | Crossref Full Text | Google Scholar
3. Zhang F, Wei K, Slowikowski K, Fonseka CY, Rao DA, Kelly S, et al. Defining inflammatory cell states in rheumatoid arthritis joint synovial tissues by integrating single-cell transcriptomics and mass cytometry. Nat Immunol. (2019) 20:928–42. doi: 10.1038/s41590-019-0378-1
PubMed Abstract | Crossref Full Text | Google Scholar
4. Rao DA, Gurish MF, Marshall JL, Slowikowski K, Fonseka CY, Liu Y, et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature. (2017) 542:110–4. doi: 10.1038/nature20810
PubMed Abstract | Crossref Full Text | Google Scholar
5. Sokolove J, Schiff M, Fleischmann R, Weinblatt ME, Connolly SE, Johnsen A, et al. Impact of baseline anti-cyclic citrullinated peptide-2 antibody concentration on efficacy outcomes following treatment with subcutaneous abatacept or adalimumab: 2-year results from the AMPLE trial. Ann Rheum Dis. (2016) 75:709–14. doi: 10.1136/annrheumdis-2015-207942
PubMed Abstract | Crossref Full Text | Google Scholar
6. Rigby W, Buckner JH, Louis Bridges S, Nys M, Gao S, Polinsky M, et al. HLA-DRB1 risk alleles for RA are associated with differential clinical responsiveness to abatacept and adalimumab: data from a head-to-head, randomized, single-blind study in autoantibody-positive early RA. Arthritis Res Ther. (2021) 23:245. doi: 10.1186/s13075-021-02607-7
PubMed Abstract | Crossref Full Text | Google Scholar
7. Platt AM, Gibson VB, Patakas A, Benson RA, Nadler SG, Brewer JM, et al. Abatacept limits breach of self-tolerance in a murine model of arthritis via effects on the generation of T follicular helper cells. J Immunol. (2010) 185:1558–67. doi: 10.4049/jimmunol.1001311
PubMed Abstract | Crossref Full Text | Google Scholar
8. Verstappen GM, Meiners PM, Corneth OBJ, Visser A, Arends S, Abdulahad WH, et al. Attenuation of follicular helper T cell-dependent B cell hyperactivity by abatacept treatment in primary sjögren’s syndrome. Arthritis Rheumatol. (2017) 69:1850–61. doi: 10.1002/art.40165
PubMed Abstract | Crossref Full Text | Google Scholar
9. Glatigny S, Höllbacher B, Motley SJ, Tan C, Hundhausen C, Buckner JH, et al. Abatacept targets T follicular helper and regulatory T cells, disrupting molecular pathways that regulate their proliferation and maintenance. J Immunol. (2019) 202:1373–82. doi: 10.4049/jimmunol.1801425
PubMed Abstract | Crossref Full Text | Google Scholar
10. Scarsi M, Paolini L, Ricotta D, Pedrini A, Piantoni S, Caimi L, et al. Abatacept reduces levels of switched memory B cells, autoantibodies, and immunoglobulins in patients with rheumatoid arthritis. J Rheumatol. (2014) 41:666–72. doi: 10.3899/jrheum.130905
PubMed Abstract | Crossref Full Text | Google Scholar
11. Jenks SA, Cashman KS, Zumaquero E, Marigorta UM, Patel AV, Wang X, et al. Distinct effector B cells induced by unregulated toll-like receptor 7 contribute to pathogenic responses in systemic lupus erythematosus. Immunity. (2018) 49:725–39. doi: 10.1016/j.immuni.2018.08.015
PubMed Abstract | Crossref Full Text | Google Scholar
12. Sanz I, Wei C, Jenks SA, Cashman KS, Tipton C, Woodruff MC, et al. Challenges and opportunities for consistent classification of human B cell and plasma cell populations. Front Immunol. (2019) 10:2458/full. doi: 10.3389/fimmu.2019.02458/full
PubMed Abstract | Crossref Full Text | Google Scholar
13. Rubtsov AV, Rubtsova K, Kappler JW, Jacobelli J, Friedman RS, Marrack P. CD11c-expressing B cells are located at the T cell B cell border in spleen and are potent antigen presenting cells. J Immunol. (2015) 195:71–9. doi: 10.4049/jimmunol.1500055
PubMed Abstract | Crossref Full Text | Google Scholar
15. Bugatti S, Manzo A, Benaglio F, Klersy C, Vitolo B, Todoerti M, et al. Serum levels of CXCL13 are associated with ultrasonographic synovitis and predict power Doppler persistence in early rheumatoid arthritis treated with non-biological disease-modifying anti-rheumatic drugs. Arthritis Res Ther. (2012) 14:R34. doi: 10.1186/ar3742
PubMed Abstract | Crossref Full Text | Google Scholar
16. Chomarat P, Briolay J, Banchereau J, Miossec P. Increased production of soluble CD23 in rheumatoid arthritis, and its regulation by interleukin-4. Arthritis Rheumatol. (1993) 36:234–42. doi: 10.1002/art.1780360215
PubMed Abstract | Crossref Full Text | Google Scholar
18. Shoda H, Nagafuchi Y, Tsuchida Y, Sakurai K, Sumitomo S, Fujio K, et al. Increased serum concentrations of IL-1 beta, IL-21 and Th17 cells in overweight patients with rheumatoid arthritis. Arthritis Res Ther. (2017) 19:111. doi: 10.1186/s13075-017-1308-y
PubMed Abstract | Crossref Full Text | Google Scholar
19. Jones JD, Hamilton BJ, Challener GJ, de Brum-Fernandes AJ, Cossette P, Liang P, et al. Serum C-X-C motif chemokine 13 is elevated in early and established rheumatoid arthritis and correlates with rheumatoid factor levels. Arthritis Res Ther. (2014) 16:R103. doi: 10.1186/ar4552
PubMed Abstract | Crossref Full Text | Google Scholar
20. Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham CO, et al. 2010 Rheumatoid arthritis classification criteria: an American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. (2010) 62:2569–81. doi: 10.1002/art.27584
PubMed Abstract | Crossref Full Text | Google Scholar
21. Zhang F, Jonsson AH, Nathan A, Millard N, Curtis M, Xiao Q, et al. Deconstruction of rheumatoid arthritis synovium defines inflammatory subtypes. Nature. (2023) 623:616–24. doi: 10.1038/s41586-023-06708-y
PubMed Abstract | Crossref Full Text | Google Scholar
22. Wan B, Nie H, Liu A, Feng G, He D, Xu R, et al. Aberrant regulation of synovial T cell activation by soluble costimulatory molecules in rheumatoid arthritis. J Immunol. (2006) 177:8844–50. doi: 10.4049/jimmunol.177.12.8844
PubMed Abstract | Crossref Full Text | Google Scholar
23. Rioja I, Hughes FJ, Sharp CH, Warnock LC, Montgomery DS, Akil M, et al. Potential novel biomarkers of disease activity in rheumatoid arthritis patients: CXCL13, CCL23, transforming growth factor alpha, tumor necrosis factor receptor superfamily member 9, and macrophage colony-stimulating factor. Arthritis Rheumatol. (2008) 58:2257–67. doi: 10.1002/art.23667
留言 (0)