
記住我


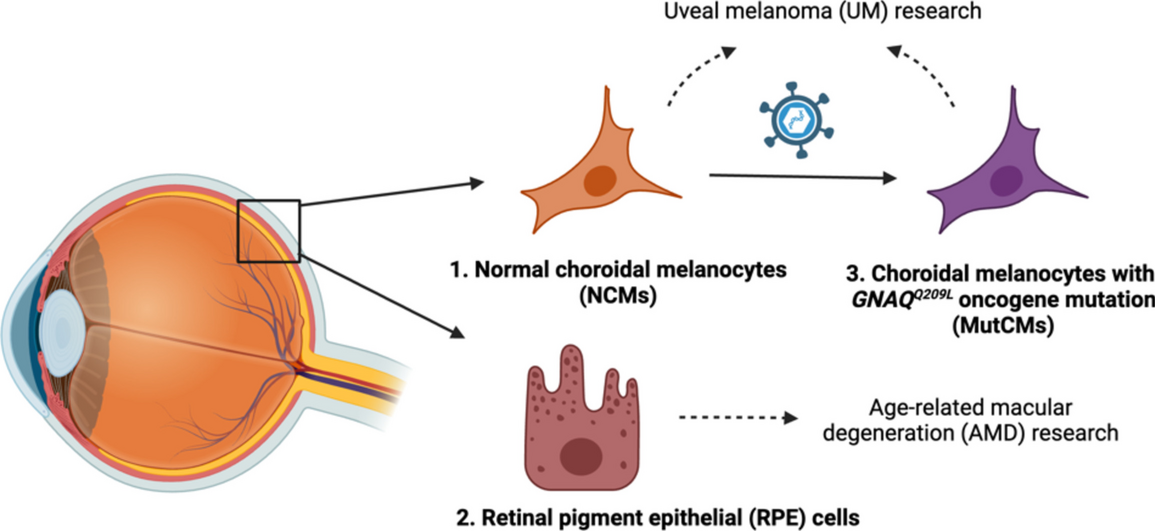



In this study, we validated a protocol for the sequential co-isolation of these cell types from single human donor eyes (Fig. 1a, b and Table 1) [19]. NCMs expressed MLANA, vimentin, HMB45, TYRP1, and S100 and were negative for the RPE markers (i.e., cytokeratin 18 (KRT18)) indicating a pure NCM culture (Fig. 1c). In parallel, isolated RPE cells expressed specific phenotypic markers (i.e., KRT18, RPE65) and were negative for the melanocyte marker HMB45, indicating a pure RPE cell culture (Fig. 1c).
Fig. 1
NCMs and RPE cells were successfully co-isolated and cultured. a An illustration of human eye anatomy and the location of RPE cells and NCMs. b Images of eye dissection and sequential RPE cell and NCM isolation. c Immunofluorescence analyses validated cell identity and purity. Scale bars = 30 μm
Table 1 Summary of donor information. Clinical information (age, sex, cause of death), days between death to cell isolation, and cell cumulative population doubling (cpd) times of NCMs and RPE cells. f: female, m: male. n.d.: not determined. NCM cpd with †: cpd of NCM culture estimated based on the splitting ratio during passages and the last passage in culture. The cultures were used up for experiments before proliferation ceased (indicated by “ > ”) or deliberately terminated due to ceased proliferation before reaching confluence (indicated by “ < ”). RPE cell cpd with †: cells analyzed as previously published [37]In total, we obtained eyeballs from 20 deceased human donors with an age range of 64 to 85 years (72.8 ± 6.2 years, 11 men and 9 women; Table 1). The success rate of viable cultures was 19/20 (95%) and 15/20 (75%) for NCMs and RPE cells, respectively. On average, over 1 to 3 months of cell culture, NCMs doubled 7.3 ± 3.6 times (n = 18 recorded cultures), while RPE cells doubled 15.2 ± 7.9 times (n = 9 recorded cultures) (Fig. 2a–d and Table 1). Days from death to cell isolation ranged from 2 to 6 days (4 ± 1.1 days). This, as well as donor sex, did not affect cell recovery and the success of cell culture (Additional file 1a-d). The donor age did not affect the cumulative population doubling (cpd) times of RPE cell cultures (P = 0.4670) but was significantly inversely correlated to the cpd of NCM cultures (P = 0.0303) (Fig. 2e, f).
Fig. 2
Characterization of the growth potential of NCMs and RPE cells. Cpd versus cultured days of a NCMs from 7 donors and b RPE cells from 4 donors, from day of isolation to end passage where proliferation ceased, observed by significant cell detachment. Cpd of all characterized c NCMs (n = 18) and d RPE cells (n = 9) are shown as the mean ± SD. e Cpd of NCMs significantly correlated to donor age (P = 0.0303). f Cpd of RPE cells did not significantly correlate to donor age (P > 0.05). g Pigment and morphology change of NCMs and RPE cells over 2 months. Cells eventually stained positive (blue) for β-galactosidase. Scale bars = 100 μm. d, days
Within 10 days of culture, both NCMs and RPE cells grew as a monolayer of adherent, brown-pigmented cells, displaying characteristic shapes (Fig. 2g). Morphologically, NCMs were pigmented and had various morphologies (bipolar, tripolar, or dendritic). RPE cells presented an epithelioid shape and were bigger in size with darker pigmentation. Both cell types gradually lost their pigmentation, reached senescence (stained positive for β-galactosidase), and ceased proliferating in around 2 months (Fig. 2g).
Since the recovery rate of NCMs from freeze–thaw was rarely reported, we analyzed the recovery rate and cpd of 14 batches of NCMs from 9 donors stored in liquid nitrogen (Additional file 1e-g). Total frozen days ranged from 9 to 618 days. Over half of the batches showed poor attachment after thawing (9/14, 64.29%) and a negative increment of cpd (8/14, 57.14%). The maximum cpd reached by freeze-thawed cells was significantly reduced compared to non-cryopreserved cells (P = 0.0028). This suggests that NCMs are highly susceptible to damage by the freeze–thaw cycle.
Together, these data confirm that NCMs and RPE cells can be efficiently co-isolated from a single eye and maintained in culture long enough to perform subsequent studies. Since we previously studied the behavior of primary RPE cells under blue light exposure [37], we focused in the present study on deciphering the best conditions for NCM maintenance and subsequent mutagenesis for the study of UM.
Serum and TPA were not necessary for NCM cultureNCMs exhibited lower growth potential when compared to RPE cells in general. Therefore, we aimed to optimize the NCM culture by comparing the morphology and confluence change in different media and the metabolic activity on different substrates.
Both serum and 12-O-tetradecanoylphorbol-13-acetate (TPA) are known to stimulate NCM growth [38, 39], but how they compare to serum-free and phorbol ester-free M2 medium has not been reported to our knowledge. We tested NCM growth of 4 different donors in 3 types of media: (1) DMEM/F12 + 10% FBS, (2) DMEM/F12 + 10% FBS + 100 nM TPA, and (3) a serum-free M2 medium. After 7 days of exposure to different media, NCMs exhibited apparent differences in cell morphology (Fig. 3a). While we observed donor-to-donor differences in confluence change, NCMs cultured in M2 medium exhibited the best growth rate consistently (Fig. 3b). Surprisingly, supplementation of TPA to DMEM/F12 + 10% FBS only improved the growth of 1/4 (25%) cultures (i.e., NCM713). In summary, serum and TPA were not necessary or insufficient to stimulate NCM growth, and NCMs responded to these two factors variably.
Fig. 3
Different cell culture media were assessed to optimize NCM culture. a Morphology of NCMs in different media on day 7. Scale bars = 100 μm. b Confluence change (%) of NCMs cultured compared to day 0. a, b n = 4 biological replicates (NCMs from 4 different donors), with each repeated in n = 4 wells. Data are shown as mean ± SD. Two-way ANOVA with Dunnett’s multiple comparisons tests (baseline: DMEM/F12 + 10% FBS) at each time point. ns: not significant. *, **, ***, and **** indicate P < 0.05, 0.01, 0.001, or 0.0001, respectively
The addition of a basement membrane matrix altered NCM growth patternsA basement membrane matrix is a specialized extracellular matrix (ECM) containing collagen IV, laminins, heparan sulfate proteoglycans, and a variety of growth factors [40]. It provides physiological culture conditions and has been used to enhance cell differentiation and survival of primary cells. It was reported that Matrigel™, a commercial basement matrix, promotes neural, melanocytic, and chondrogenic differentiation of trunk neural crest cells [41]. In addition, the thicknesses of the Matrigel™ layer and cell seeding density have been shown to affect the cell morphology of breast metastatic cells [40]. The effect of Matrigel™ on melanocyte morphology and survival has not been previously reported. To understand whether Matrigel™ could stimulate the NCM growth, we first tried culturing a late passage of NCM114 in a 24-well plate coated with 80 μL Matrigel™ per well or on uncoated wells (Fig. 4a). While the NCMs constantly detached on non-coated wells over the 6 days, they did not detach on Matrigel™-coated wells, and they migrated to form spheroids. Based on this observation, we next sought to study the effect of Matrigel™ on NCM cultures more comprehensively by coating wells with different matrix thicknesses and using different cell seeding densities in 96-well plates.
Fig. 4
Different cell culture substrates were assessed for optimizing NCM culture. a Morphology of NCM114 (late passage, P4) seeded on a 24-well plate coated without or with 80 μL of Matrigel™ per well at 18,333 cells per well. Scale bars = 100 μm. b Morphology of NCMs on different substrates. In a 96-well plate, 2500 cells were seeded per well and were imaged after 3 days. Scale bars = 100 μm. c The metabolic activity of NCMs was assessed after 3 days of culture on different substrates. Data are shown as mean ± SD. One-way ANOVA with multiple comparisons test (baseline: no coating), ns: not significant. **P < 0.01, ****P < 0.0001. d Morphology of NCMs on Matrigel™ at various cell seeding densities. In a 96-well plate coated with 45 μL of Matrigel™ per well, 1000, 3000, 5000, or 7000 cells were seeded per well and were imaged after 7 days. b–d n = 4 biological replicates (NCMs from 4 different donors), with each repeated in n = 3 wells. Scale bars = 400 μm
We tested NCM growth on different amounts of Matrigel™ (15 μL, 30 μL, and 45 μL per well of a 96-well plate) and a cell attachment enhancer, poly-l-lysine. The poly-l-lysine-coated surface is positively charged and thus enhances the electrostatic interaction between the negatively charged cell membrane and the surface. Morphologically, NCMs cultured without coating presented typical melanocyte shapes (bi/tripolar and dendritic), while they were more flattened and rounded on poly-l-lysine (Fig. 4b). With the increasing thickness of Matrigel™, the dendrites of NCMs became more retracted. On 45 μL of Matrigel™, NCMs exhibited mostly rounded morphology. After 3 days of culture, a CCK-8 assay was performed to analyze cell metabolic activity, an index for cell vitality (Fig. 4c). Surprisingly, the metabolic activity was not enhanced by the addition of poly-l-lysine or Matrigel™ coating but was inhibited in 3/4 (75%) cultures (P < 0.01).
Lastly, we tested different cell seeding densities on a thick layer of Matrigel™ (45 μL per well) to determine if the confluency would impact cell growth characteristics. Interestingly, over 7 days of culture, NCMs remained mostly as single cells when seeded at low density (1000 or 3000 cells per well), while spheroids were seen when seeded at higher density (5000 or 7000 cells per well) (Fig. 4d). Based on daily observation, the spheroids appeared to result from cell migration, instead of growth in foci. Further study on this phenomenon is warranted. In summary, although donor-to-donor variations were seen, Matrigel™ did not enhance NCM metabolic activity but altered cell morphology and growth behavior.
GNAQ Q209L MutCMs were generated successfully with high mutant GNAQ copiesGNAQ/11Q209L mutation is believed to be a major initial event for UM oncogenesis. However, the generation of mutant premalignant choroidal melanocytes of human origin has not been previously reported. The efficient isolation of NCMs has prompted us to generate a GNAQQ209L MutCMs from CMs that would serve as a powerful model to study UM oncogenesis and development.
As NCMs are precious cells that are limited in quantity, we practiced and validated the lentiviral transduction method on multiple helper cell types including BJ fibroblasts and HEK293 (Additional file 2a-b). These cell types are easy to grow and were used for testing the virus preparation infectivity before its use on NCMs. A viral dose-dependent mutant copy fraction was found, and the freshly prepared virus was more infectious than the freeze-thawed virus. In addition, polybrene, a cationic polymer, was a critical factor in enhancing the transduction efficiency (Additional file 2c).
After validating the virus infectivity on other cell types, we sought to test it on NCMs. Four NCM donors were transduced at the same time under the same condition (50,000 cells were mixed with 50 μl of fresh virus in 1 ml of M2 medium supplemented with 4 μg/ml polybrene). Consistently, all NCMs were efficiently transduced with high mutant copies (0.9783 ± 0.0081 fraction of all GNAQ copies being mutant, Fig. 5a, b). Successful introduction of the point mutation (GNAQ (c.626A > T)) was further validated by Sanger sequencing (Additional file 3). MutCMs with high mutant copies (mutant fraction > 0.8) were heterogenous in morphology, with a dendritic shape and long processes (Fig. 5a, b, Additional file 2d-e). MutCMs expressed MLANA, vimentin, HMB45, TYRP1, and S100 and were negative for KRT18 as expected (Fig. 5c).
Fig. 5
The insertion of the mutant gene (GNAQ (c.626A > T)) by lentiviral transduction was highly efficient. a Phase contrast images and ddPCR 2D plots for MutCMs. b The transduction efficiency, indicated by wildtype (WT) and mutant (Mut) fraction. c MutCMs (MutCM713 was shown) expressed MLANA, Vimentin, HMB45, TYRP1, and S100 and were negative for KRT18. Scale bar = 30 μm. d The fraction of WT and Mut GNAQ/11 gene in 4 UM cell lines (92.1, MP46, MEL270, and MP41) were profiled. e Representative images and digital droplet PCR (ddPCR) of NCMs and MutCMs and f their WT and Mut copy fraction. Scale bars = 100 μm. ddPCR plots: droplets that are positive for WT, Mut, and both gene copies are shown in green, blue, and orange, respectively. Negative droplets are shown in black
To compare to the mutant GNAQ/11 mutant fraction in UM cell lines, we analyzed the wildtype (WT) and GNAQ/11 mutant (Mut) copies in 4 human UM cell lines: 92.1 and MP46 (GNAQ (c.626A > T)), MEL270 (GNAQ (c.626A > C)) and MP41 (GNA11 (c.626A > T)). Mutant fractions ranged from 0.3225 to 0.5789 (Fig. 5d). Therefore, we believed the mutant copy fraction similar to this range may be more relevant to the context of UM. By titrating various virus dosages on NCMs (early passage, P < 3) or helper cell lines in order to tune the virus dosage, 6 MutCMs were successfully transduced with mutant fractions ranging from 0.2095 to 0.6296 (Fig. 5e, f, Additional file 2d-e). Interestingly, compared to their parental NCMs, these MutCMs tended to grow in clusters instead of a monolayer (Fig. 5g, Additional file 2e). The level of clustering and spheroid formation varied between donors.
In addition to the lentiviral transduction method for the genetic engineering of NCMs, we also explored electroporation for future genome editing applications such as creating relevant melanocyte mutants using CRISPR-Cas9. We tested the electroporation efficiency of an EGFP plasmid in NCMs and UM cells (Additional file 4). Optimal electroporation conditions differed between NCMs (1400 V, 20 ms, 2 pulses) and UM cells (1300 V, 20 ms, 1 pulse) (Additional file 4a) as well as the percentage of obtained fluorescent cells (48.9% and 83.5%, respectively) (Additional file 4b). Electroporating NCMs could thus be a valuable technique in various research fields related to vision science research.
In summary, the described methods were highly efficient for establishing MutCMs. Further characterization of MutCMs is warranted to understand behavioral and molecular changes that may recapitulate the formation of nevi.
Proteomic profiling of NCMs, MutCMs and UMsTo further our understanding of premalignant CM models, we sought to compare proteomic profiles of NCMs (NCM602, 713, 809, 1009) and UMs (92.1, MP46, MP41). Secondly, we compared a pair derived from the same donor (NCM713 and MutCM713). Tandem mass tag (TMT)-mass spectrometry identified 3268 proteins after quality control of the data. All cells expressed melanocyte markers (e.g., MLANA, VIM, PMEL, TYRP1, and S100B) with higher levels in NCMs and MutCMs compared to UM cell lines (Fig. 6a). NCMs (n = 4) and UMs (n = 3) showed robust difference in the volcano plot (Fig. 6b), with 549 and 191 proteins being significantly downregulated and upregulated, respectively (P < 0.05, fold change < 0.5 or > 2). Gene ontology (GO) term analysis revealed distinct enrichment of biological processes, molecular functions, and cellular components in NCMs and UMs. Notably, ATP metabolic process, oxidation–reduction, cell–matrix adhesion, and cell morphogenesis involved in differentiation were upregulated in NCMs, while mRNA metabolic process, RNA splicing, and gene expression were upregulated in UMs (Fig. 6c). The Reactome pathway enrichment analysis is shown in Table 2. In concordance with GO terms, NCM proteome profiles were enriched in pathways such as tricarboxylic citric acid (TCA) cycle and respiratory electron transport, cell-extracellular matrix interactions, cell junction organization, and biological oxidation. In addition, innate immune system and apoptosis pathways are elevated in NCMs. UM proteome profiles were enriched in pathways such as metabolism of RNA, translation, mRNA splicing, and regulation of TP53 activity, which highly aligns with the findings of a recently published study [42].
Fig. 6
Proteomic comparisons of NCMs and UMs. a Heatmap of relative expression of melanocyte markers shown in z-score. b Volcano plot of proteins in NCMs vs. UMs. Significantly upregulated and downregulated proteins in NCMs and UMs appear in blue and red, respectively (P < 0.05, Log2(fold change) < − 1 or > 1). c Gene ontology analysis of the significantly different proteins. Selected terms are with P < 0.05, Benjamini < 0.05. A full list can be found in Additional file 6
Table 2 Selected upregulated pathways in NCMs and UM cells, respectively (P < 0.05). A full list can be found in Additional file 6In contrast, differences between NCM713 and MutCM713 were subtle (Table 3), with only 36 proteins being differentially regulated (P < 0.05, fold change < 0.67 or > 1.5) (Table 3). Interestingly, 5 out of 8 downregulated proteins in MutCM713 are involved in mitochondrion, and 2 downregulated proteins (i.e., STK10, COMMD5) are thought to have tumor suppressor properties [43, 44]. Upregulated proteins in MutCM713 include ALDH1A1, which is a cancer stem cell marker [45] and has been found overexpressed in melanoma cells [46]. CASP3 and UBE2E3 are involved in regulating apoptosis and were elevated. Importantly, numerous upregulated proteins in MutCM713 have been implicated in cancer progression, including PAK4 [47], MKI67 [48], PSMD10 [49], ELP3 [50, 51], and STAT6 [52]. In summary, the proteomic changes in MutCM713 may provide novel insights and understanding of UM oncogenesis.
Table 3 Differentially expressed proteins in NCM713 and MutCM713 (P < 0.05, fold change < 0.67 or > 1.5)Transcriptomic profiling of NCMs, MutCMs, and UMsTo validate the proteomic data, we performed RNA sequencing on NCMs and MutCMs derived from the same donor (NCM713, MutCM713) and a UM cell line (92.1). Over 26,000 identities of RNA with unique official gene symbols were identified after quality control. Consistent with the proteomic results, NCM713 and MutCM713 have generally higher expression levels of melanocyte markers (i.e., MLANA, VIM, PMEL, TYRP1, S100B, MITF, DCT, SOX10, EDNRB, GPNMB) compared to 92.1 (Fig. 7a). Differential gene expression was analyzed and a P value < 0.05 and Log2(fold change) < − 1 or > 1 was considered significant. Specifically, for NCM713 vs. 92.1, 3536 and 2918 genes are downregulated and upregulated in 92.1, respectively (Fig. 7b). NCM713 and MutCM713 have fewer differences, as expected, with 1541 and 1957 genes downregulated and upregulated in MutCM713, respectively (Fig. 7c). Five hundred eighteen and 700 genes are commonly downregulated and upregulated in 92.1 and MutCM713, respectively (Fig. 7d).
Fig. 7
Transcriptomic comparisons of NCM713, MutCM713, and 92.1. a Heatmap of relative expression of melanocyte markers shown in z-score. Volcano plot of b NCM713 vs. 92.1 and c NCM713 vs. MutCM713. Significantly upregulated and downregulated genes appear in blue and red, respectively (P < 0.05, Log2(fold change) < − 1 or > 1). d Venn diagram of differential gene expression analysis of NCM713 vs. 92.1 and NCM713 vs. MutCM713 (P < 0.05, Log2(fold change) < − 1 or > 1). e Selected regulated pathways identified by KEGG Pathway Enrichment Analysis of commonly regulated genes in 92.1 and MutCM713. A full list can be found in Additional file 6. f Heatmap of selected gene expression of interest
To understand whether MutCM713 shares cancer-associated malignant properties with 92.1, the commonly significant genes were queried against KEGG Databases (Fig. 7e, f). Top enriched pathways of downregulated genes include cell adhesion molecules, motor proteins, cell cycle, gap junctions, and p53 signaling pathway, with reduced expression of genes such as CDH1, CDH4, ITGA6, TP53I3, AIFM2, and CHECK2. The top 3 enriched pathways of upregulated genes are pathways in cancer, PI3K-Akt signaling pathway, and MAPK signaling pathway, followed by other prominent signaling associated with cell survival, migration, and aggressiveness, including Ras, WNT, Rap1, Hippo, Notch, and TGF-beta signaling pathways. Notably, upregulated gene expression include PLCB4, ERBB4, RASGRP3, TRAF6, SOS1, PIK3CA, AKT3, MYC, WNT4, WNT11, WNT88, VEGFA, VEGFC, JAG2, and NOTCH3. Complete lists of differentially expressed genes and significant KEGG pathways have been provided in the Additional file 6. Collectively, transcriptomic profiling data provides further insights into the potential molecular changes in GNAQQ209L-mutated pre-cancer cell models (MutC713) and UM cells (92.1).
Overview of current culture methods and research applications using human NCMsThere is a lack of literature review on melanocyte culture methodologies, especially within the context of ocular research. Considering this gap, we summarized the current methodologies for establishing ocular cell cultures and research applications, focusing particularly on human NCMs, that are not commercially available (Fig. 8). This can serve as a reference guide of validated protocols and techniques according to downstream analyses.
Fig. 8
Overview of current methods in generating NCMs and RPE cells, and NCM culture methods and research applications. Created in https://BioRender.com. MSH, melanocyte-stimulating hormone; bFGF, basic fibroblast growth factor; cAMP, cyclic adenosine monophosphate
The isolation procedures include eyeball dissection and sequential enzymatic dissociation aimed at isolating NCMs and RPE cells. Notably, Hu et al. compared the use of trypsin, collagenase, or both enzymes for the isolation of melanocytes from the choroidal stroma [18]. Among these methods, a trypsin treatment followed by an incubation in collagenase produced the highest number of viable NCMs. Contamination by RPE cells and fibroblasts was eliminated by Geneticin. Another significant development in this arena was by Valtink and Engelmann, who described a protocol for the co-isolation of RPE cells and NCMs [19, 53]. The authors observed that collagenase treatment for one hour was sufficient in dissociating RPE cells but not for NCMs, allowing for sequential isolation of RPE cells followed by NCMs using dispase. Apart from the conventional ex vivo extraction techniques, multiple studies have created RPE cells from induced pluripotent stem cells [20]. Lastly, oncogene-mutated melanocytes, which mimic melanoma precursor cells, have been created from human and murine skin melanocytes [8, 54], but to the best of our knowledge, nothing similar has been documented for NCMs. We therefore propose the methodologies for creating MutCMs by lentiviral transduction or electroporation for genome editing.
While RPE cells can be easily grown in an FBS-supplemented medium [20], the culture of NCMs requires specific growth and melanin-stimulating agents [18, 39, 55]. Hu et al. suggested that a melanocyte culture medium requires three classes of supplements, such as serum, mitogens (e.g., phorbol esters or bFGF), and cAMP enhancers (e.g., cholera toxin, isobutyl methylxanthine) [39, 55]. Phorbol esters were found to be mitogenic and melanin-stimulating [39, 55]. Basic FGF, insulin, and cAMP enhancers were shown to be important for melanocyte proliferation [39, 55]. Some neurotransmitters could influence melanocyte growth and melanogenesis [56]. Stem cell factor was mitogenic in the presence of certain essential factors [57]. Although melanocyte-stimulating hormone was proven to induce melanocyte growth and melanogenesis for skin melanocytes, its effect on NCMs was controversial [33, 58].
As crucial as the culture medium, the substrate could influence melanocyte survival and morphology. Thereby, the choices of culture conditions rely on the research objectives and applications. Research on NCMs mainly focused on melanin biogenesis [24, 55, 56, 59,60,61,62], oncogenesis (primarily as a control for studying UM) [63,64,65,66,67,68,69,70,71,72,73,
留言 (0)