
記住我


A summary of the pathological features of 244 PDAC and 236 TNBC samples is given in Supplementary Table 1. The IHC staining of MET and RON was performed in the paired tumor tissue using specific antibodies. The immunoreactive intensity, patterns, and positivity were evaluated by referencing the FDA-approved criteria for EGFR positivity [44]. Representative images of MET and RON were obtained using the negative to positive staining technique with different intensities (Fig. 1a, b). The individual PDAC and TNBC samples exhibited similar patterns of MET and RON expressions, but with heterogeneous appearances. Both samples exhibited predominantly membrane, predominantly cytoplasmic, and mixed staining patterns (Fig. 1a, b), suggesting differential expression patterns of MET and RON in a single tumor mass. Hence, we can suggest that MET and RON were differentially expressed in the PDAC and TNBC samples with different staining intensities and patterns, reflecting the heterogeneous nature of individual cancer cells.
Fig. 1: Immunohistochemical staining and analysis of MET and RON expression in primary PDAC and TNBC samples.
a Differential expression of MET and RON in 244 primary PDAC samples. b Differential expression of MET and RON in 236 TNBC samples. IHC staining of MET and RON using their corresponding specific antibodies, respectively, was performed as detailed in Materials and Methods. MET and RON expression levels were determined using semi-quantitative methods [14]. Staining intensities of MET and RON were determined as negative, weak, moderate, or strongly positive. The area reactivity of staining was set as 0 = < 1%, 1 = 2%–30%, 2 = 31%–70%, and 3 = 71%–100%. The combined scores that define MET and RON expressions were as follows: 0, negative; 1–2, low; 3–4, moderate; and 5–6, high, corresponding to the scale of 0, 1+, 2+, and 3+, respectively. Various patterns of MET or RON expression with different IHC staining intensities were observed. The representative images are presented. IHC immunohistochemistry, MET mesenchymal-epithelial transition, RON recepteur d’Origine nantais, PDAC pancreatic ductal adenocarcinoma, TNBC triple negative breast cancer.
A summary of MET and RON expression in 244 PDAC samples is given in Table 1. A total of 218 and 225 PDAC samples expressed MET (89.35%) and RON (92.21%), respectively. Tumors from 117 PDCA patients showed low-to-moderate levels of MET expression (53.67%). Similarly, 136 cases had low-to-moderate RON expression levels (60.44%). Overexpression was observed in 101 cases for MET (46.33%) and 89 cases for RON (39.56%). Among the 218 cases showing MET positivity, 157 cases exhibited co-expression with RON (72.02%). Similarly, in the 225 cases demonstrating RON positivity, 141 exhibited co-expressed RON (62.67%). Matched co-overexpression was observed only in 23.39%–22.67% of the total number of cases for both MET and RON.
Table 1 Immunohistochemical analysis of MET and RON expression by immunohistochemical staining in primary pancreatic ductal adenocarcinoma and triple negative breast cancer samplesa.Among the 236 TNBC samples, 198 samples expressed MET (83.90%) and 185 samples expressed RON (78.37%) (Table 1). Tumors from 108 patients showed low-to-moderate levels of MET expression (55.45%). Similarly, 94 cases had low-to-moderate levels of RON expression (50.81%). Overexpression was observed in 90 cases for MET (45.46%) and 91 cases for RON (49.19%). Among the total 198 MET-positive cases, 127 co-expressed RON (64.14%), while among the 185 RON-positive cases, 134 exhibited MET expression. The matched co-overexpression in both cases was 30.30% and 32.42%, respectively (Table 1). These findings indicate that MET and RON are co-expressed in variable combinations at high frequencies, suggesting that MET and RON expressional heterogeneity is a pathogenic feature in both pancreatic and breast cancers.
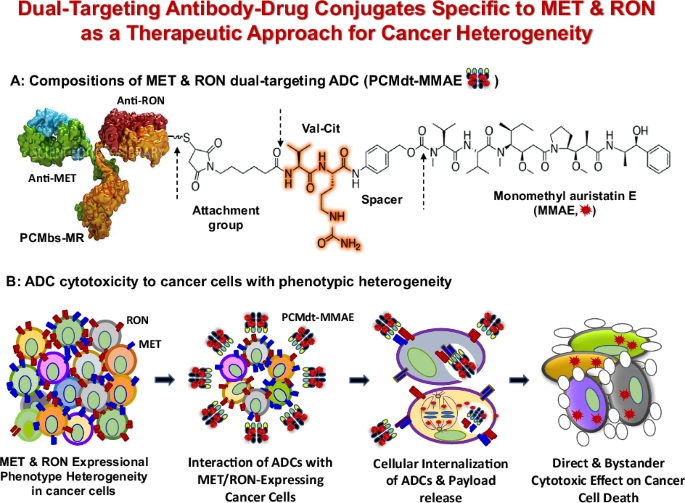
Generation of MET and RON bispecific mAbs and dual targeting ADCsAnti-MET mAb PCM-MET01 and anti-RON mAb PCM-5B14 were selected to construct PCMbs-MR—the humanized IgG1 bispecific antibody (Fig. 2a, c). PCMbs-MR was appropriately modified and optimized to ensure the proper formation of bispecific IgG molecules. The resulting PCMbs-MR was then utilized to generate MET and RON dual-targeting ADC PCMdt–MMAE with a DAR of 4:1 (Fig. 2b). PCMbs–MR was specifically validated to interact with cell-surface MET only, RON only, and both MET and RON (Fig. 2d). PCMbs–MR still exhibits considerable binding capacity in tumor cell lines expressing RON or MET alone. We analyzed the species-specificity of PCMbs–MR and found that it binds to MET and RON only from humans and monkeys, not to those from canines or mice (Fig. 2e). It exhibits a combination-binding affinity of 0.836 µg/mL (5.57 nM) for human MET and RON and that of 0.914 µg/mL (6.09 nM) for monkey MET and RON homologs.
Fig. 2: Generation and characterization of bispecific monoclonal antibody PCMbs-MR.
a Schematic structure of PCMbs-MR generated through cDNA recombination from anti-MET mAb PCM-MET01 and anti-RON mAb PCM5B14. A single heavy chain and light chain, representing both anti-MET mAb and anti-RON mAb, respectively, is presented. The knob-into-hole was created in the CH3 domains of both anti-MET and anti-RON mAbs [42]. The crossover was conducted between the CH1 domain of the heavy chain and the CL domain of the light chain from the anti-MET mAb accordingly [43]. b PCMbs-MR is conjugated with MMAE through a cleavable dipeptide linker to form the dual-targeting ADC PCMdt-MMAE [32,33,34]. The drug-to-antibody ratio (DAR) was calculated as 4.13:1. c Structural 3D analysis of individual complementary-determining regions (black) grafted into the variable regions of humanized heavy chain and light chain (top panel). Antigen-binding surfaces in the variable regions of the humanized anti-MET and anti-RON mAbs are depicted as dark blue (bottom panel). d Cell surface immunofluorescent analysis of PCMbs-MR binding to MET and RON. A panel of cancer cell lines expressing variable levels of MET, RON, or both receptors were used. Anti-MET mAb PCM-MET01 and anti-RON mAb PCM5B14 were included for comparison. Briefly, cells at 1 × 106 cells per sample were incubated with 2 µg/mL of PCMbs-MR or other antibodies (red line), followed by rabbit anti-human or mouse IgG coupled with FITC. Regular human IgG was used as the control (black line). Immunofluorescence intensities from individual samples were analyzed using a flow cytometer [32]. Cellular levels of receptor expressions are indicated as follows: HCC1806(MET-/RON-); Hs746T(MET3+/RON-); SUM52PE(MET-/RON3+); MDA-MB468(MET1+/RON2+); HT29(MET3+/RON3+); BxPC-3(MET3+/RON2+); T-47D(MET3+/RON3+); HCC1937(MET3+/RON3+); HCC2185(MET3+/RON-); and MDA-MB-231(MET3+/RON2+). e Species specificity of PCMbs-MR. Human HT29, monkey 4MBr-5, canine MDCK, and mouse MS-1 cell lines, known to express both MET and RON, were used. Incubation of cells with PCMbs-MR and specific immunofluorescence analysis were performed as detailed previously [32]. The binding affinity of PCMbs-MR was calculated using the GraphPad Prism 6 software. f PCMbs-MR induced cell-surface MET and RON internalization by individual cancer cells. Seven cancer cell lines expressing variable levels of MET, RON, or both receptors were used. PCMbs-MR at 5 µg/mL was used for incubation. Procedures used to determine cell surface receptor internalization were performed as previously described [32,33,34]. Internalization efficacy (IE50) among cell lines was calculated using a previously described method [32,33,34]. g PCMbs-MR-induced internalization efficacy in comparison with anti-MET or anti-RON mAbs. BxPC-3 and HCT116 cell lines expressing high levels of both MET and RON were used. Treatment of cells with mAbs and the methods used to determine IE50 were carried out to obtain the individual IE50 values [32]. ADC antibody–drug conjugate, FITC fluorescein isothiocyanate, mAb monoclonal antibody, MET mesenchymal-epithelial transition, MMAE monomethyl auristatin E, PCMbs–MR humanized bispecific monoclonal antibody specific to both MET and RON, RON recepteur d’Origine nantais.
The ability of PCMbs-MR to induce cell-surface MET, RON, and both receptor internalization is shown in Fig. 2f. A panel of cell lines with different levels of MET and RON and their combinational expression patterns were used. The average time required for PCMbs–MR to induce a 50% reduction in the MET and RON receptor levels (IE50) ranged from 10.21 to 22.86 h. Importantly, PCMbs-MR was found to be effective in inducing the internalization of both MET and RON, as well as their receptors in cell lines exhibiting heterogeneous expression patterns (Fig. 2d–f). A comparison of PCMbs–MR with anti-MET and anti-RON mAbs in receptor internalization using the PDAC cell line BxPC-3(MET3+/RON2+) and colorectal cancer cell line HCT116(MET2+/RON3+), both of which co-express high levels of MET and RON, is shown in Fig. 2g. The average IE50 value for PCMbs–MR (9.68 h) is comparable to those of PCM–MET01 (10.85 h) and PCM5B14 (12.66 h). Thus, PCMbs–MR is specific to both MET and RON. As bispecific binding causes a robust internalization of cell-surface MET and RON by cancer cells, PCMbs–MR is an ideal candidate for targeted drug delivery.
Pharmacokinetic profiles and toxic effects of PCMdt–MMAEA single-dose injection of 10 or 30 mg/kg of PCMdt–MMAE was injected into mice, with or without BxPC-3 cell-mediated xenograft tumors. The objective was to determine whether the PK profile of PCMdt–MMAE could be altered in mice bearing tumors expressing MET and RON. Mice without tumors were used to determine the target-independent behavior of PCMdt–MMAE. The PK profile of PCMdt–MMAE in the plasma of both tumor-bearing and tumor-nonbearing mice appeared to exhibit the two-compartment model with similar patterns for both low and high doses (Fig. 3a, b). In tumor-bearing mice, PCMdt–MMAE had an average mean plasma clearance of 0.20 mL·d−1·kg−1, a t½ of 6.33 days, and a mean residential period of 6.95 days (Fig. 3a). These values were similar to those in mice without tumors (0.21 mL·d−1·kg−1, 6.19 days, and 7.07 days, respectively) (Fig. 3b). Overall, data from tumor-bearing mice overlapped with those from tumor-nonbearing mice with 95% prediction intervals. Thus, the presence of tumors with MET and RON expression does not affect the dynamics of PCMdt–MMAE.
Fig. 3: Pharmacokinetic profiles and toxic activities of PCMdt-MMAE in mice.
Analysis of PCMdt-MMAE PK profiles: Athymic nude mice (8-week-old) were divided into tumor-bearing (a) and tumor-nonbearing (b) groups (three animals per group). Mice from the tumor-bearing group were subcutaneously injected with 1 × 106 BxPC-3 cells. When tumor volumes reached ~500 mm3, both groups of mice were administered once through the tail vein with 3 or 10 mg/kg of PCMdt-MMAE. Blood samples were collected from individual mice at different time intervals. The amount of MMAE-conjugated PCMbs-MR in plasma was determined by using a MMAE ADC ELISA kit (Eagle Biosciences Inc., Nashua, NH, USA). The PK parameters were calculated using the WinNonlin software package (Certara, Princeton, NJ, USA) [32,33,34]. c Toxic effects of PCMdt-MMAE in vivo: Effects of multiple doses of PCMdt-MMAE on mouse bodyweight were determined by a single administration of PCMdt-MMAE at 10, 30, and 60 mg/kg, respectively. The mice were weighed and monitored for 12 days. The average bodyweight before PCMdt-MMAE injection was 19.8 ± 3.6 g (five mice per group) and set as 100%. ADC, antibody–drug conjugate; MMAE, monomethyl auristatin E; PCMdt-MMAE, monomethyl auristatin E was conjugated to PCMbs–MR to generate the dual-targeting ADC; PCMbs-MR humanized bispecific monoclonal antibody specific to both MET and RON, PK pharmacokinetic
To determine the maximum tolerated dose of PCMdt–MMAE, the mice were categorized into three groups based on the single-dose injection of PCMdt–MMAE (10, 30, and 60 mg/kg) (Fig. 3c). Mice injected with 10 mg/kg of PCMdt–MMAE behaved normally during the observation period. The average body weight of the experimental group was comparable to that of control mice, with no significant differences. Slightly distressed activity was observed in mice administered with 30 mg/kg of PCMdt–MMAE. These mice also showed an average reduction of ~5% (P > 0.05 in comparison with control mice) in body weight within the first 5 days of PCMdt–MMAE injection. This reduction was, however, recovered on day 6 of the observation period. In contrast, a 20% reduction in body weight was observed in mice treated with 60 mg/kg of PCMdt–MMAE (P < 0.05, compared with control mice). The overall bodyweight of these mice remained slightly lower than that of control mice, with a difference of ~3% on day 12. Thus, a single dose of <30 mg/kg of PCMdt–MMAE appears to be relatively safe without any statistical differences in the activity, behavior, or body weight of the animals.
Toxicity profiles were studied using Sprague–Dawley rats with a single-dose injection of PCMdt–MMAE (10 and 30 mg/kg). All animals survived at the end of the study period (28 days). No gross abnormal changes were observed in the daily activity, bodyweight, food consumption, or responsiveness of the animals. However, abnormalities were observed in the hematology and blood workup of the mice (Supplementary Table 2). Notably, 30 mg/kg, not 10 mg/kg, of PCMdt–MMAE caused moderate reductions in leukocytes, as evident by the decrease in the populations of neutrophils, lymphocytes, and monocytes. Blood chemistry analysis revealed varying levels of the increase in blood enzymatic activities, including those of alanine transaminase, alkaline phosphatase, aspartate aminotransferase, and creatine kinase, indicating damage to the liver and other tissues. In addition, moderate myelosuppression in the bone marrow was observed with 30 mg/kg of PCMdt–MMAE. These adverse reactions explain the effects of MMAE [32]. All abnormalities were temporary, reversible, and normalized to baseline levels by the end of the study, suggesting that up to 30 mg/kg of PCMdt–MMAE can be safely used.
Effects of PCMdt–MMAE on cancer cell cycle, viability, and deathThe pathogenic features of cancer cell lines are presented in Supplementary Table 3. The effects of PCMdt–MMAE on the cell cycle were studied using BxPC-3 cells. HCC1806 cells were employed as the control. PCMdt–MMAE treatment of BxPC-3 cells led to dramatic changes in their cell cycles (Fig. 4a). This effect could be observed as early as 12 h after the addition of the ADC and was characterized by a significant reduction in the G0/G1 phase, a decrease in the S phase, and an increase in the G2/M phase. As expected, HCC1806 cells did not respond to PCMdt–MMAE. Quantitative measurements of cell cycle changes are shown in Supplementary Table 4. These results suggest that PCMdt–MMAE has a profound impact on the cell cycle of cancer cells heterogeneously co-expressing MET and RON.
Fig. 4: Biological effects of PCMdt-MMAE on CRC cell cycle, survival, and death.
a Changes in cell cycle: BxPC-3 and control HCC1806 (1 × 106 cells per dish) cells were treated at 37 °C with 5 µg/mL of PCMdt-MMAE for various times. They were then collected, stained with propidium iodide, and analyzed using a flow cytometer [32,33,34]. Changes in cell cycle were marked with arrows. b Reduction of cell viability: A panel of seven cancer cell lines expressing variable levels of MET, RON, or both receptors (5000 or 8000 cells per well in a 96-well plate in triplicate) were treated with different amounts of PCMdt-MMAE for 96 h. HCC1806 cells without MET or RON expression served as the control. Cell viability was determined by the MTS assay [32]. c Dose-dependent cell death: Three cancer cell lines expressing MET, RON, or both receptors were treated with different amounts of PCMdt-MMAE for 96 h as described in (b). HCC1806 cells were used as the control. At the end of the study, the dead cells were counted using the Trypan blue exclusion assay to determine the percentage of cell death [32]. d Morphological evidence of cell death. Treatment of cells with PCMdt-MMAE was performed as described in (c). Cellular morphological changes from individual cell lines were observed at 96 h under the Olympus BK-41 inverted microscope and photographed. For all studies described above, the percentages of cell viability and/or cell death and the individual IC50 values from individual groups were calculated using the GraphPad Prism 6 software. Results shown here are from one of three experiments with similar results. ADC antibody–drug conjugate, CRC colorectal cancer, MET mesenchymal-epithelial transition, PCMdt-MMAE monomethyl auristatin E was conjugated to PCMbs–MR to generate the dual-targeting ADC, RON recepteur d’Origine nantais
The effect of PCMdt–MMAE on cell viability was studied using six cancer cell lines (HCC1806, BxPC-3, HT29, SUM52PE, Hs746T, and ASPC-1) expressing varying levels of MET and RON, as well as their heterogeneous combinations. PCMdt–MMAE treatment in a dose-dependent manner led to a significant reduction in cell viability (Fig. 4b). The IC50 value of PCMdt–MMAE at 96 h was within 1.34–3.78 μg/mL, with an average value of 2.52 μg/mL. A comparison of the ability of PCMdt–MMAE to reduce cancer cell viability with that of anti-MET and anti-RON ADCs is shown in Supplementary Table 5. The average IC50 values of PCMdt–MMAE were comparable to those of anti-MET and anti-RON ADCs. Thus, PCMdt–MMAE is effective not only in cancer cells expressing MET, RON, and their heterogeneous combinations at high levels, but also in cells co-expressing both MET and RON at relatively low levels.
We further determined the effect of PCMdt–MMAE on cell death using three cancer cell lines (Hs746T, SUM52PE, and BxPC-3) expressing MET, RON, and their heterogeneous combinations. PCMdt–MMAE can kill cancer cells in a dose-dependent manner, with IC50 values of 2.91 µg/mL ± 0.91 for Hs746T(MET3+/RON-), 3.90 µg/mL ± 0.40 for SUM52PE(MET-/RON3+), and 1.55 µg/mL ± 0.08 for BxPC-3(MET3+/RON2+) (Fig. 4c). The morphological observation indicated cell death on a massive scale 96 h after the cells were exposed to PCMdt–MMAE (Fig. 4d), although cellular sensitivities to PCMdt–MMAE-induced cell death varied among the three cell lines tested. Thus, based on the results depicted in Fig. 4c, d, along with those in Fig. 4a, b, it can be suggested that PCMdt–MMAE is active not only in cancer cells with only MET or RON expression, but also in those co-expressing both receptors. The effect of PCMdt–MMAE appears to be manifested through cell cycle arrest, cell viability reduction, and large-scale cell death.
Therapeutic activity of PCMdt–MMAE against tumors heterogeneously co-expressing MET and RONThe first objective of our study was to determine the efficacy of PCMdt-MMAE using PCM-MET01-MMAE and PCM5B14-MMAE as references. Tumors initiated by HT29 colon cancer cells(MET3+/RON3+) were used. Individual ADCs were administered in a single dose of 10 mg/kg to tumor-bearing mice. The growth of HT29 cell-mediated tumors was dramatically inhibited among all three groups of mice treated with individual ADCs (Fig. 5a and Table 2A). PCMdt-MMAE exhibited long-lasting antitumor activities up to day 32 without any sign of tumor regrowth. The efficacy of PCMdt–MMAE, as determined by using its tumoristatic concentration (TSC), is comparable to that of anti-MET and anti-RON ADCs without any statistical significance (Fig. 5a).
Fig. 5: Therapeutic efficacy of PCMdt-MMAE in xenograft tumor models.
Mice bearing xenograft tumors were divided into experimental and control groups (five animals per group). ADCs were injected through the tail vein. RhIgG-MMAE was used as the control. Tumor volumes were measured every four days. Individual tumors were collected at the end of the study, photographed, weighed to reach an average value per group, and analyzed for statistical differences using GraphPad Prism 6 software. The tumoristatic concentration (TSC), a minimal dose required to maintain a balance between the growth and inhibition of xenograft tumors, was calculated [32,33,34]. a Therapeutic efficacy of PCMdt-MMAE in comparison with anti-MET or anti-RON ADCs. HT29 cell-mediated xenograft tumors were used. PCMdt-MMAE, PCM-MET01-MMAE, and PCM5B14-MMAE at 10 mg/kg were injected once into mice. Mice treated with RhIgG-MMAE served as the control. Tumor growth was monitored up to day 36 or 40. b, c Dose-dependent effects of PCMdt-MMAE on xenograft tumor growth. FG and T-47D cell-derived xenograft tumors were used. PCMdt-MMAE from 1, 3, 7, 10, and 15 mg/kg in a Q12×2 schedule was injected once into mice. d Duration of PCMdt-MMAE-mediated anticancer activity. Xenograft tumors derived from H358, BxPC-3, and HCT116 cell lines were used. HCC1806 tumors served as the control. PCMdt-MMAE at 10 mg/kg was used for treatment. Tumor growth from each group was monitored until tumor regrowth was visible. ADC antibody–drug conjugate, MET mesenchymal-epithelial transition, PCMdt-MMAE monomethyl auristatin E was conjugated to PCMbs–MR to generate the dual-targeting ADC, RON recepteur d’Origine nantais.
Table 2 A Therapeutic efficacy of PCMdt-MMAE at a single dose of 10 mg/kg in comparison with that of anti-MET ADC PCM-MET-1-MMAE or anti-RON ADC PCM5B14-MMAE against tumor xenografts mediated by colorectal cancer HT29 cellsa. B Dose-dependent efficacy of PCMdt-MMAE against tumor xenografts mediated by PDAC FG and breast cancer T-47D cells in mouse modelsb. C Therapeutic effect of PCMdt-MMAE on multiple tumor xenografts mediated by T-47D, FG, HT29, and BxPC-3 cell lines in mouse modelsc.Our second objective was to determine the dose-dependent activity of PCMdt–MMAE (Fig. 5b, c, and Table 2B). T-47D(MET2+/RON3+) and FG(MET3+/RON2+) cell lines heterogeneously co-expressing MET and RON were utilized. PCMdt–MMAE, which inhibited both FG and T47-D cell-derived tumor growth in a dose-dependent manner, was administered at doses of 1, 3, 7, 10, and 15 mg/kg in a Q12×2 schedule. Notably, tumor growth could be delayed to up to day 28 or 32 with 1 mg/kg of PCMdt–MMAE (Fig. 5b, c, and Table 2B). Long-term inhibition of tumor growth was noted with 7, 10, and 15 mg/kg of PCMdt–MMAE (Fig. 5b, c, and Table 2B). In these cases, no sign of tumor regrowth was observed until day 44. A further analysis of the average tumor number and tumor weight at the end of the study period demonstrated that PCMdt–MMAE reduced tumor weights in a dose-dependent manner (Table 2B). The T-47D tumors exhibited a significant reduction in tumor weight, ranging from 65% to 99%, depending on the PCMdt–MMAE dose. Moreover, PCMdt–MMAE, at all administered doses, was able to eradicate tumors to varying levels (Table 2B). Thus, PCMdt–MMAE is highly effective against tumors heterogeneously co-expressing both MET and RON.
Our final objective was to determine the duration of PCMdt–MMAE-mediated activity. The H358(MET2+/RON3+), HCT116(MET2+/RON3+), and BxPC-3(MET3+/RON2+) cells showing varying levels of MET and RON co-expressions were used. A single-injection regimen of PCMdt–MMAE comprised a dose of 10 mg/kg. The growth of tumors caused by all the aforementioned three cancer cell lines was inhibited upon PCMdt–MMAE administration (Fig. 5d and Table 2C). The average TSC value of PCMdt–MMAE was 0.29 mg/kg for these tumors. Signs of tumor regrowth among the three models tested were observed only around day 36 or 40, equivalent to six t½ (~6.3 days) cycles of PCMdt–MMAE. An analysis of the average tumor volume and weight at the end of the study indicated that PCMdt–MMAE is highly effective in inhibiting tumor growth. Average tumor volume reductions of 86.42%, 97.00%, and 90.90% were observed for H358 at day 36, HCT116 at day 28, and BxPC-3 at day 32, respectively. Notably, PCMdt–MMAE exhibited variable tumor-eradicating activities (Table 2C). Thus, a single-dose regimen of 10 mg/kg of PCMdt–MMAE is highly effective with long-lasting activity against tumors initiated by the aforementioned three cancer cell types with varying levels of MET and RON co-expressions.
留言 (0)