
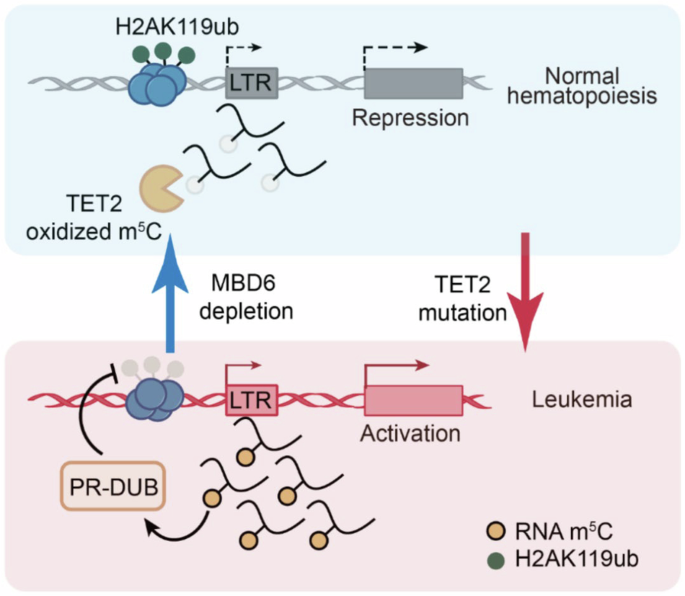
記住我
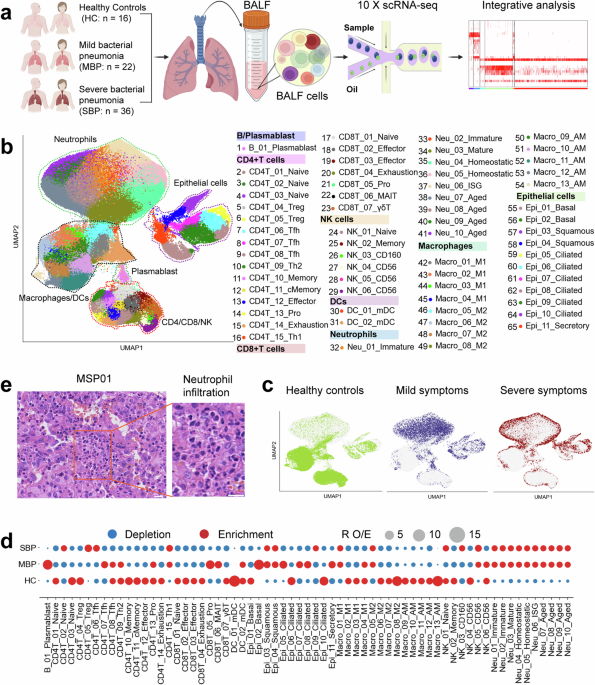

To characterize the immune landscape in bacterial pneumonia, single-cell transcriptomic profiles were generated from bronchoalveolar lavage fluid (BALF) samples obtained from 58 patients and 16 healthy controls (HCs) using droplet-based scRNA-seq (10x Genomics platform) (Fig. 1a). The 58 patients diagnosed with bacterial pneumonia were categorized into two clinical groups: mild bacterial pneumonia (MBP, n = 22) and severe bacterial pneumonia (SBP, n = 36). The majority of causative agents are prevalent bacterial pneumonia pathogens, such as Acinetobacter baumannii, Pseudomonas aeruginosa, and Staphylococcus aureus, among others. Detailed clinical information and laboratory findings of the 58 enrolled patients are provided in Supplemental Table 1 and Supplementary Fig. 1. We obtained 444,146 high-quality single cells, with an average of 3500 unique molecular identifiers (UMIs) representing 1164 genes (Supplementary Fig. 2a–d). Of these cells, 60,084 cells (13.5%) were from the HC condition, 94,590 cells (21.3%) were from the MBP condition, and 289,472 cells (65.2%) were from the SBP condition (Supplementary Fig. S2a–d). After adjusting for read depth and mitochondrial read counts, all high-quality cells were merged into an unbatched and comparable dataset for subsequent principal component analysis (See methods).
Using graph-based clustering of uniform manifold approximation and projection (UMAP), we identified transcriptomic profiles for 8 major cell types: plasmablasts, macrophages, neutrophils, CD4+ T, CD8+ T, natural killer (NK) cells, dendritic cells (DCs), and epithelial cells, based on the expression of canonical gene markers (Fig. 1b and Supplementary Fig. 2e). In the UMAP space, lymphoid cells (T and NK) were clearly differentiated from macrophages and neutrophils, whereas epithelial cells presented a distinct transcriptomic pattern from that of immune cells (Fig. 1b and Supplementary Fig. 2f). When colored by disease-level, the UMAP representation were segregated by severity following bacterial infection, particularly in severe patients compared to healthy controls, indicating bacterial infection-induced transcriptomic changes (Fig. 1c and Supplementary Fig. 3a). We firstly identified 45 cell subtypes within 8 major cell clusters (Supplementary Fig. 2f). Further subclustering revealed a total of 65 subtypes encompassing diverse respiratory cell types (Fig. 1b and Supplementary Fig. 4), which was used for subsequent analyses. Consequently, we clearly defined the cell subpopulation compositions in BALFs, providing an information-rich dataset for accurate annotation and comprehensive analysis of these cell types at different resolutions.
Each cell cluster displayed different sample origins and enrichment in disease severities (Fig. 1c and Supplementary Fig. 3). To analyze the unique immune profiles of different disease groups, we investigated the immune cell composition in each individual (Fig. 1d and Supplementary Fig. 3). Notably, the percentages of neutrophils in BALFs were significantly increased, particularly in patients with severe bacterial pneumonia (Fig. 1d, e and Supplementary Fig. 3e).17 In contrast to the elevation of neutrophils in BALFs, most other major immune cell types decreased in patients with bacterial pneumonia (Fig. 1d and Supplementary Fig. 3). Importantly, the preference of each cell subtype in different patient groups was also illustrated according to RO/E: the ratio of observed to expected cell frequencies (employed to mitigate the influence of technical variation on disease-associated cellular composition),18 with proliferating B (B_01_Plasmablast), CD4+ T (CD4T_13_Pro) and CD8+ T (CD8T_05_Pro) cells more enriched in mild patients compared to severe cases and healthy controls (Fig. 1d). The plasma B cells (B_01_Plasmablast) in BALFs exhibited high expression of genes encoding the constant regions of immunoglobulin G1 (IgG1), IgG3, IgG4, or IgGM (Supplementary Fig. 3f), suggesting their function in secreting antigen-specific antibodies. These findings indicate that effective B cell activation and antibody production may play an important role in bacterial pneumonia. Overall, the data revealed that each disease state in patients with bacterial pneumonia might exhibit a unique immunological fingerprint.
Macrophage and neutrophils are the main drivers of the lung inflammatory response in patients with bacterial pneumoniaThe host’s inflammatory response against the causative microorganism is crucial for resolving bacterial pneumonia.19 To investigate the potential origins of cytokine production, we established a cytokine score and an inflammatory score for each immune cell by analyzing the expression of cytokine genes and known inflammatory response genes, respectively (Supplementary Table 2).20 Cytokine and inflammatory scores were defined using the sc.tl.score_genes function in Scanpy.12 Hyperinflammatory cell populations were identified by performing pairwise comparisons of these scores across all subtypes using Kruskal–Wallis tests with Bonferroni correction. These two scores were utilized as indicators to evaluate the potential inflammatory impact of different immune cell populations. In patients with bacterial pneumonia, we observed significant upregulation of cytokine and inflammatory genes, indicative of an inflammatory response (Fig. 2a and Supplementary Fig. 5a). Patients with severe bacterial pneumonia exhibited significantly higher cytokine and inflammatory scores than patients with mild bacterial pneumonia, indicating the presence of an inflammatory cytokine storm in lungs of severely affected patients (Supplementary Fig. 5a).
Fig. 2
Myeloid cells are the primary contributors to the production of pro-inflammatory cytokines in severe patients. a UMAP projections of BALFs. Colored based on the eight major cell types (top left), eight hyper-inflammatory cell subtypes (top right), cytokine score (Middle), and inflammatory score (Bottom). b Pie charts depicting the relative contribution of each inflammatory cell subtype to the cytokine and inflammatory scores in severe patients. c Heatmap depicting the expression of cytokines within each hyper-inflammatory cell subtype identified. d Lollipop chart depicting the relative contribution of the top ten cytokines in patients with severe disease. e Box plots of cytokine expression based on scRNA-seq and plasma profiling for healthy controls, mild patients, and severe patients. Significance was evaluated using the Kruskal–Wallis test with Bonferroni correction significance was evaluated using the Kruskal–Wallis test with Bonferroni correction (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, nsp > 0.05). f Heatmap plots of the sum of significant interaction among the eight hyper-inflammatory cell subtypes. g Dot plot of the interactions among inflammatory macrophages in severe patients. P values are indicated by the circle sizes, as shown in the scale on the right
Fourteen cell subtypes, including six subtypes of neutrophils, seven subtypes of macrophages, and one subtype of epithelial cells, exhibited significantly elevated cytokine and inflammatory scores based on our scRNA-seq data for BALFs (Supplementary Fig. 5b), suggesting their potential roles as sources of the inflammatory storm. Further analysis identified eight inflammatory cell clusters, including three macrophage clusters (Macro_02_M1, Macro_04_M1 and Macro_10_AM) and five neutrophil clusters (Neu_01_Immature, Neu_02_Immature, Neu_04_Homeostatic, Neu_05_Homeostatic, Neu_06_ISG), which exhibited significantly higher cytokine and inflammatory scores in severe patients compared to those with mild patients and healthy donors (Fig. 2a and Supplementary Fig. S6). These findings suggest that these cell clusters are likely the primary contributors to the cytokine storm observed in severe patients. We further investigated the relative abundance of these eight cell subsets in severe patients and observed a notable increase, further implicating them in the heightened inflammatory response seen in severe cases (Supplementary Fig. 5c).
Subsequent analysis revealed that four inflammatory subsets, including Macro_02_M1, Neu_01_Immature, Neu_04_Homeostatic, and Neu_05_Homeostatic, contributed over 80% of the inflammatory and cytokine scores (Fig. 2b), indicating that these cells were the predominant drivers of inflammation in severe patients. This finding aligns with previous studies on other infectious diseases such as COVID-19 where macrophages and neutrophils were also identified as key drivers of cytokine storms.12,13 Analysis of inflammatory signatures within the identified subsets revealed distinct pro-inflammatory cytokine gene expression profiles for each inflammatory cell cluster (Fig. 2c), including CXCL1/2/3/8, S100A8/9/12, IL1RN, PPBP, etc. Moreover, we observed elevated expression of key inflammatory cytokines, including CXCL1/2/3 and S100A8/9/12, in patients with severe symptoms (Supplementary Fig. 7a). These findings suggest that diverse mechanisms may drive the inflammatory storm in severe pneumonia. The elevated expression of cell type-specific pro-inflammatory cytokines in four main subsets of inflammatory cells (Fig. 2c and Supplementary Fig. 7b) further validates their core role in driving the inflammatory storm seen in those with severe disease.
Ten pro-inflammatory cytokines (CXCL1/2/3/8, S100A8/9/12, IL1RN, FGF13 and TNFSF12), which are primarily expressed in Macro_02_M1, Neu_01_Immature, Neu_04_Homeostatic and Neu_05_homeostatic (Supplementary Fig. 7c), are critical drivers of the inflammatory storm, accounting for over >99% of cytokine scores in severe patients (Fig. 2d). S100A8/A9 and CXCL8 were the most significant contributors, comprising approximately 92% of cytokine scores (Supplementary Fig. 2d). Patients with severe disease exhibited elevated expressions of S100A8/A9 and CXCL8 genes (Fig. 2e), further supporting our hypothesis. BALF analysis within our cohort revealed significantly higher concentrations of S100A8/A9 complex and CXCL8 (also known as IL-8) in severe patients, corroborating our scRNA-seq findings (Fig. 2e). This underscores the significance of the hyper-inflammatory cell subsets and molecules (CXCL8 and S100A8/A9) as possible therapeutic targets for mitigating the immunopathogenesis seen in severely infected pneumonia patients.
The cytokine storm seen in severe patients may result from complex cellular crosstalk among hyperinflammatory cell subtypes, mediated by the release of various cytokines.13 To further investigate this phenomenon, we examined the ligand-receptor interaction patterns among eight distinct hyperinflammatory cell subsets in these patients (Fig. 2f and Supplementary Fig. 7e, f). Our analysis revealed several noteworthy ligand-receptor interactions within these subtypes, particularly highlighting the heightened interactions among inflammatory macrophages (Fig. 2f and Supplementary Fig. 7e). These inflammatory macrophages expressed multiple receptors, including SLC7A1, CCR5, CCR2, IL15RA, HAVCR2, etc., suggesting responsiveness to various cytokines secreted by other cells (Fig. 2g and Supplementary Fig. 7f). Notably, our analysis confirmed that the interactions between inflammatory macrophages and other hyperinflammatory cells clusters primarily rely on chemokines and their corresponding receptors (Fig. 2g and Supplementary Fig. 7f). These findings provide molecular insights into the potential mechanisms underlying complex network of interactions between hyperinflammatory cell subsets in severe pneumonia.
Tfh and Th2 cell expansion observed in mild patients but not in severe patientsA total of 15 CD4+ T cell subtypes were identified, including three naïve (CD4T_01_Naive, CD4T_02_Naive, and CD4T_03_Naive), two regulatory (CD4T_04_Treg and CD4T_05_Treg), three follicular helper (CD4T_06_Tfh, CD4T_07_Tfh, and CD4T_08_Tfh), Th2 (CD4T_09_Th2), memory (CD4T_10_Memory), central memory (CD4T_11_cMemory), proliferating (CD4T_13_Pro), exhausted (CD4T_14_Exhaustion) and Th1 (CD4T_15_Th1) T cells (Fig. 3a). Analysis of disease preference revealed that Tfh, Th2, and proliferating CD4+T cells were enriched in mild patients, while a Treg subset (CD4_05_Treg) expanded predominantly in severe patients (Fig. 1d), suggesting that a Tfh and Th2 response might be involved in controlling the bacterial infection.
Fig. 3
Immunological features of CD4+ T cell subsets. a The clustering result (Left row) of 15 CD4+ T cell types (right row) from 74 samples. Each point represents one single cell, colored according to the CD4+ T cell subtype. b Correlation between plasmablast with Tfh cells in mild patients. c Dot plots showing the expression of selected genes in Th2 cells across disease conditions. d UMAPs illustrating IFN-I response and unhelped signature scores for each CD4+ T cell. The red circle highlights the CD4T_14_Exhaustion cluster, which is characterized by high expression of interferon-I (IFN-I) response genes and an unhelped T cell signature. e Flow cytometry plots showing gating strategy and typical exhausted molecules in CD8+T cells from bacterial pneumonia (Top row: severe bacterial pneumonia; Bottom row: mild bacterial pneumonia). f Dot plots showing the cell exhaustion-related markers in CD4_13_Pro and CD4T_14_Exhaustion across disease conditions. g Venn diagram illustrating the number of upregulated genes in CD4+ T cells. h Enriched GO biological process terms for upregulated genes in CD4+ T cells from mild (Left) and severe (Right) disease. Only select terms are shown. i Dot plots showing CD4+ T cells related genes across disease conditions
CD4T_06/07/08_Tfh cells exhibited a transcriptional profile consistent with T follicular helper (Tfh) cells (e.g., high expression of inducible costimulatory; ICOS) (Supplementary Fig. 4), indicating their potential function in facilitating a protective B cell immune response. Tfh cells are crucial for generating optimal antibody responses and ensuring the production of high-quality neutralizing antibodies during pathogen infection.21 Consistent with this, we also observed a notable expansion of plasmablasts in individuals with mild symptoms (Fig. 1d), along with a strong positive correlation between the increased Tfh cells and expanded plasmablasts in these individuals (Fig. 3b). However, this correlation diminishes in cases of increased severity (Supplementary Fig. 8a). The primary function of Th2 cells is to facilitate B cell activation, and the cytokines they secrete enhance B cell proliferation, differentiation, and antibody production.22 Interestingly, among mild patients, there was a strong correlation observed between the expanded Th2 cells and the elevated plasmablasts (Supplementary Fig. 8b). Key genes and transcription factors associated with Th2 polarization and B cell activation exhibited higher expression levels in mild patients compared to both severe patients and healthy controls (Fig. 3c). Taken together, these findings suggest a coordinated response between T cells and B cells in mild patients, leading to effective humoral immunity against bacteria, which appears to become uncoupled in severe patients.
Pathogen infection can lead to lymphocyte exhaustion.12 We, therefore, profiled the expression of genes encoding typical markers of exhaustion in CD4+ T cells. Among CD4+T clusters, two subtypes (CD4T_13_Pro and CD4T_14_Exhaustion) exhibited notably higher exhaustion scores (Supplementary Fig. 8c), with the highest exhaustion score observed in mild patients (Supplementary Fig. 8d). The lack of substantial CD4+ T cell exhaustion in severe patients (Supplementary Fig. 8d) indicates that CD4+ T cell exhaustion may not be a significant contributor to severe bacterial pneumonia. It is well-documented that the activation of T cell exhaustion is closely connected to sustained type I interferon (IFN) signaling and inadequate CD4 + T cell assistance. In mild patients, the exhaustion profile identified within the CD4T_14_Exhaustion and CD4T_13_Pro clusters appeared to be associated with constant type I IFN signaling and insufficient assistance from CD4+ T cells, as anticipated (Fig. 3d). In mild cases, exhausted CD4 + T cells consistently displayed elevated expression of inhibitory genes, including PDCD1, HAVCR2, LAG3, CTLA4, and TIGIT, along with exhaustion-associated transcriptional factors such as PTPN6/11 and PRDM1 (Fig. 3e).
To identify relevant biological functions in disease-specific upregulated genes, we performed a Gene Ontology (GO) analysis of biological pathways. In CD4+ T cells, we detected 2654 and 694 genes upregulated for mild and severe cases versus controls, respectively, while there were 638 upregulated genes in the comparison between the two disease groups (Fig. 3f and Supplementary Table 3). The abundance of differentially expressed genes (DEGs) indicated a substantial variation within CD4+ T cells among mild patients. GO terms associated with upregulated DEGs in mild bacterial pneumonia patients were primarily associated with antibacterial defense and immune protection, including neutrophil activity and antigen processing, whereas those in severely affected patients were more associated with inflammatory reactions, cell apoptosis, tissue damage, and immunosuppression (Fig. 3g). Genes (e.g., CXCL8, ID2, HSPB1, and PER1), which are associated with inflammatory reactions and immunosuppression, were found to be highly expressed in severe patients (Fig. 3h). In addition, we observed that a specific subset of Treg cells (CD4T_04_Treg) exhibited high expression levels of TGF-β in patients with severe symptoms (Supplementary Fig. 8e). Treg cells are known for their role in producing TGF-β, which helps suppress CD4+ T cells, inhibit T cell cytokine production, and downregulates immune responses from effector cells.23 Consequently, the regulatory function of Treg cells appears to be associated with the development of immune tolerance and the persistence of bacterial infections in severely affected patients. Collectively, our findings suggest that the CD4+ T cell response in patients with mild symptoms primarily supports immune protection, whereas in severe cases, it contributes to both inflammatory reactions and immunosuppression.
Activation of CD8+T cells is associated with reduced disease severity in bacterial pneumoniaSeven CD8+ T cell subclusters were identified in this study: naïve (CD8T_01_Naive), effector (CD8_02/03_Effector), exhausted (CD8_04_Exhaustion), MAIT (CD8_06_MAIT), γδT (CD8_07_γδT), and proliferating (CD8_05_Pro) (Fig. 4a). The CD8T_05_Pro subset, identified as highly proliferative, showed elevated levels of MKI67 and TYMS, along with various effector genes (e.g., NKG7, GZMA, and PRF1), supporting its classification as proliferative effector CD8+ T cells. Analysis of disease preference indicated that mucosal-associated invariant T (MAIT) cells and proliferating CD8+ T cells were predominantly found in patients with mild symptoms, whereas a depletion of all CD8+ T cell subsets was observed in patients with severe symptoms (Fig. 1d). The reduced abundance of CD8T_05_Pro cells and its progenitor cells (CD8T_02_Effector) from individuals with severe symptoms suggests some disruption to the adaptive immune responses mediated by CD8+ T cells in these cases (Fig. 1d). These findings highlight important differences in CD8+ T cell clusters between mild and severe disease bacterial pneumonia.
Fig. 4
Immunological features of CD8+ T cell subsets. a The clustering result (Left row) of seven CD8+ T cell types (right row) from 37 samples. Each point represents one single cell, colored according to cell type. b PAGA analysis of CD8+ T cell pseudo-time: the associated cell type and the corresponding status are listed. c Box plots showing the exhausted scores in CD8_04_Exhaustion and CD8_05_Pro subsets across disease conditions. Significance was evaluated using the Kruskal–Wallis test with Bonferroni correction (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, nsp > 0.05). d Dot plots showing the cell exhaustion-related markers in CD8_04_Exhaustion and CD8_05_Pro subsets across disease conditions. e Flow cytometry plots showing gating strategy and typical exhausted molecules in CD8+ T cells from patients with severe (top row) and mild (below row) bacterial pneumonia. f UMAPs illustrating IFN-I response and unhelped signature scores for each CD8+ T cell. g Dot plots showing the cytotoxicity-related genes in CD8+ T cell subsets in patients with bacterial pneumonia. h Venn diagram illustrating the number of upregulated genes in CD8+ T cells. i Dot plots showing the activation-related genes in CD8+ T cell across disease conditions
A partition-based graph abstraction (PAGA) analysis was performed to examine the developmental trajectories of the seven CD8+ T subclusters (Fig. 4b). The analysis identified the proliferating subcluster (CD8_05_Pro) and the exhausted subcluster (CD8T_04_Exhaustion) as two distinct branches. The PAGA map revealed that two nodes (CD8T_02/03_Effector) showed high connectivity, suggesting their potential role as trans-differentiation bridges among CD8+ T cell subtypes. These effector states (CD8T_02/03_Effector) appear to function as intermediate stages, linking various subtypes from naive (CD8T_01_Naive) to more activated CD8+ T cell subclusters (e.g., CD8T_04_Exhaustion and CD8T_05_Pro). Interestingly, there was significant connectivity observed between CD8T_02_Effector and CD8T_03_Effector. The developmental trajectory towards the exhaustion and proliferation subset was also observed (Fig. 4b). This trajectory correlated with functional scores in various CD8+ T cell clusters, for instance, CD8T_01_Naive and CD8T_06_MAIT had high naive scores while CD8T_04_Exhaustion and CD8T_05_Pro clusters had high exhaustion scores. The highest cytotoxic score was observed in the CD8T_05_Pro cluster while the highest inflammatory score was in the CD8T_04_Exhaustion cluster (Supplementary Fig. 9).
Two subclusters (CD8T_04_Exhaustion and CD8T_05_Pro) exhibited pronounced exhaustion states (Supplementary Fig. 9), indicating that bacterial infections may lead to CD8+ T cell exhaustion, a critical factor contributing to the imbalance in the antibacterial infection response. Interestingly, severe patients did not display higher exhaustion scores compared to those with mild symptoms, suggesting that CD8+T cell exhaustion may not be a primary factor in severe bacterial infections. Consistently, CD8T_04_Exhaustion and CD8T_05_Pro cells from patients, particularly those with mild symptoms, showed increased levels of expression from various exhaustion genes (e.g., LAG3, HAVCR2, CTLA4, and PDCD1) when compared with healthy patients (Fig. 4d). PDCD1 binds to PD-L1/PD-L2 and HAVCR2 (Tim-3) binds to galectin-9. This binding then leads to the recruitment of tyrosine-protein phosphatases SHP1 and/or SHP2 via their intracellular domains, including the immunoreceptor tyrosine-based switch motif (ITSM) and immunoreceptor tyrosine-based inhibitory motif (ITIM).13 Consequently, these interactions inhibit key signaling pathways such as PI3K-AKT and LAT-Zap70, resulting in reduced cellular proliferation and cytokine produced. Furthermore, elevated PRDM1 levels have been associated with exhausted cells having diminished polyfunctionality and increased expression of inhibitory receptors.13 Correspondingly, exhausted CD8+ T cells from individuals with bacterial infections, especially in mild cases, had notably higher expression levels of key transcription factors such as PRDM1 and PTPN6/11 compared to healthy donors (Supplementary Fig. 10a). Importantly, the CD8+ T cell exhausted phenotypes from patients with bacterial pneumonia were confirmed with flow cytometry (Fig. 4e and Supplementary Fig. 11, 12).
The development of the exhaustion program in CD8+ T cells is closely associated with continuous type I interferon (IFN) signals.24 Our analysis demonstrated that exhausted CD8+ T cells had a notable enrichment of genes related to type I IFN signaling, which indicates a direct connection between the exhaustion status of CD8T_04_Exhaustion and CD8T_05 _Pro cells and persistent type I IFN signaling (Fig. 4f and Supplementary Fig. 10b). Furthermore, the exhaustion program in CD8+ T cells was also strongly related to compromised CD4+ T cell-mediated support, which helps maintain a robust CD8+ T cell immune response in infection.24 We thus analyzed whether the exhausted CD8+ T cells (CD8T_04_Exhaustion and CD8T_05 _Pro) exhibited signs of reduced CD4+ T cell help. As expected, both CD8T_04_Exhaustion and CD8T_05_Pro showed notable upregulation of transcripts typically associated with CD8+ T cells lacking adequate CD4+ T cell assistance (Fig. 4e and Supplementary Fig. 10b). Altogether, these data suggest that the exhaustion observed in the CD8T_04_Exhaustion and CD8T_05_Pro subsets may be driven by ongoing type I IFN signaling coupled with a lack of adequate CD4+ T cell assistance.
Despite exhibiting exhausted and “unhelped” features, exhausted CD8+ T cell subclusters showed obvious positive enrichment of cytotoxicity signature genes (Supplementary Fig. 9) and elevated expression levels of cytotoxic transcripts (e.g., CST7, GNLY, PRF1, GZMA/B, NKG7) (Fig. 4g), indicating that these exhausted subtypes may be heterogeneous (Supplementary Fig. 10c). These finding aligns with past reports,24,25 which has shown that while their capacity from proliferation and production of cytokines may be reduced, cytotoxic capabilities are often still intact in exhausted CD8+ T cells. Patients, especially in the mild group, displayed higher levels of cytotoxic signature gene expression in exhausted CD8+ T cells compared to healthy controls (Supplementary Fig. 10c). Similar to our findings in exhausted CD8+ T cells, we also observed notable expression of cytotoxic genes in the CD8T_07_γδT cell population (Supplementary Fig. 10c). Effector CD8+ T cells exhibit cytolytic functions enabling them to directly eliminate pathogens or infected cells through granule-mediated mechanisms involving perforin, granzyme, and granulysin.12 Therefore, the enhanced expression of various cytolytic molecules in CD8+ T cells may be associated with immune protection in patients with mild conditions. These findings suggest that CD8 cells have undergone complete activation, particularly in individuals with mild symptoms.
To examine the characteristics of CD8+ T cells in bacterial pneumonia patients further, we investigated the DEGs in individuals with mild and severe disease relative to healthy controls. In CD8+ T cells, we observed 2012 (76.2%) and 42 (1.5%) genes that were upregulated in mild and severe cases, respectively. (Fig. 4h and Supplementary Table S4). Additionally, a total of 588 (22.3%) genes were commonly upregulated in both disease groups (Fig. 4h and Supplementary Table S4). Notably, individuals with mild symptoms exhibited greater variability in the abundance of DEGs within CD8+ T cells (Fig. 4h). CD8+ T cells isolated from BALFs, in particular CD8T_04_Exhaustion and CD8T_05_Pro, highly expressed various activation markers (e.g., CD69, CD38, CD25(IL2RA) and PDCD1) in mild patients, further suggesting that CD8+ T cells in individuals with mild symptoms are characterized by a higher degree of activation (Fig. 4i). In contrast, DEGs in CD8+ T cells derived from severely ill patients exhibited elevated expression of genes linked to inflammatory response and cell apoptosis (Fig. 4i).
Dysregulated neutrophil response as a potential contributor of severe bacterial infectionThe subclustering analysis uncovered remarkable diversity within the neutrophil compartment, identifying 10 transcriptionally distinct cell subtypes. These subtypes comprised of two immature clusters, one mature cluster, four aged clusters, one interferon-stimulated genes (ISG)-related cluster, and two homeostatic clusters (Fig. 1b and Fig. 5a). All neutrophil subsets were enriched in individuals with bacterial infections (Fig. 1d, e), confirming the significant involvement of infiltrated neutrophils. This increased abundance emphasizes the crucial role of neutrophils in combatting bacterial pathogens and underscores their function as a fundamental element of the innate immune system.26 When visualizing neutrophils at the disease-level using UMAP, neutrophils grouped according to disease severity (Fig. 5b), suggesting that neutrophils respond differently to varying degrees of bacterial infection. PAGA trajectory analysis unveiled three distinct fates of neutrophils, with Neu_03_mature, Neu_06_ISG, and Neu_07_Aged representing distinct endpoints (Fig. 5c). The developmental trajectory was correlated with cellular maturation, interferon response, and cellular senescence (Fig. 5c), implying that neutrophils may undergo distinct developmental and functional changes after bacterial infection. These findings revealed that neutrophils exhibit varying responses depending on the severity of the bacterial infection.
Fig. 5
Immunological features of neutrophils. a The clustering result (Left row) of neutrophil subtypes (right row) from 74 samples. Each point represents one single cell, colored according to the neutrophil subtype. b UMAP plot of neutrophil clusters showing disease distribution. c PAGA analysis of neutrophil pseudo-time: the associated cell type and the corresponding status are listed. d Heatmap plots of selected genes in immature neutrophil subset across disease conditions. e Violin plots of S100A8, S100A9, and S100A12 in immature neutrophil subset across disease conditions. f Heatmap plots of the sum of significant interaction between immature neutrophil clusters (Neu_01/02_Immature) and T cell subtypes in severe patients. g Venn diagram illustrating the number of upregulated genes in neutrophils. h Enriched GO biological process terms for upregulated genes in neutrophils from severe disease. Only select terms are shown. i Box plots showing the functional scores in neutrophils across disease conditions. Significance was evaluated using the Kruskal–Wallis test with Bonferroni correction (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, nsp > 0.05)
Neutrophils obtained from BALF samples of patients with bacterial infections were found to contain two distinct clusters of LDNs (low-density neutrophils), designated as Neu_01_Immature and Neu_02_Immature (Fig. 5a). LDNs are primarily generated during illness (e.g., severe infection and sepsis during emergency myelopoiesis) and are strongly associated with impaired immune reactions that exhibit both immunosuppression and inflammation.27 Immature neutrophils derived from patients with severe bacterial infections highly expressed a range of pro-inflammatory molecules, such as S100A8/9/12, CCL3/4, IL1B, and CXCL8 (Fig. 5d, e), which have been identified as key inducers of inflammatory cytokine storms in severe infectious diseases like tuberculosis and COVID-19.13 Immature neutrophils exhibited elevated expression of PADI4 (Fig. 5d), a crucial factor implicated in NETs (NETosis; neutrophil extracellular trap formation). Recent studies have demonstrated the significant involvement of NETs in the pathogenesis of severe infectious diseases.28 In addition, immature neutrophils were found to express the BPI gene, which has previously been associated with adverse outcomes in severe infections.27 Arginase 1 (ARG1), recognized as an inhibitor of T cell activation,27 was also found to be highly expressed in immature neutrophils from patients with severe bacterial pne
留言 (0)