
記住我
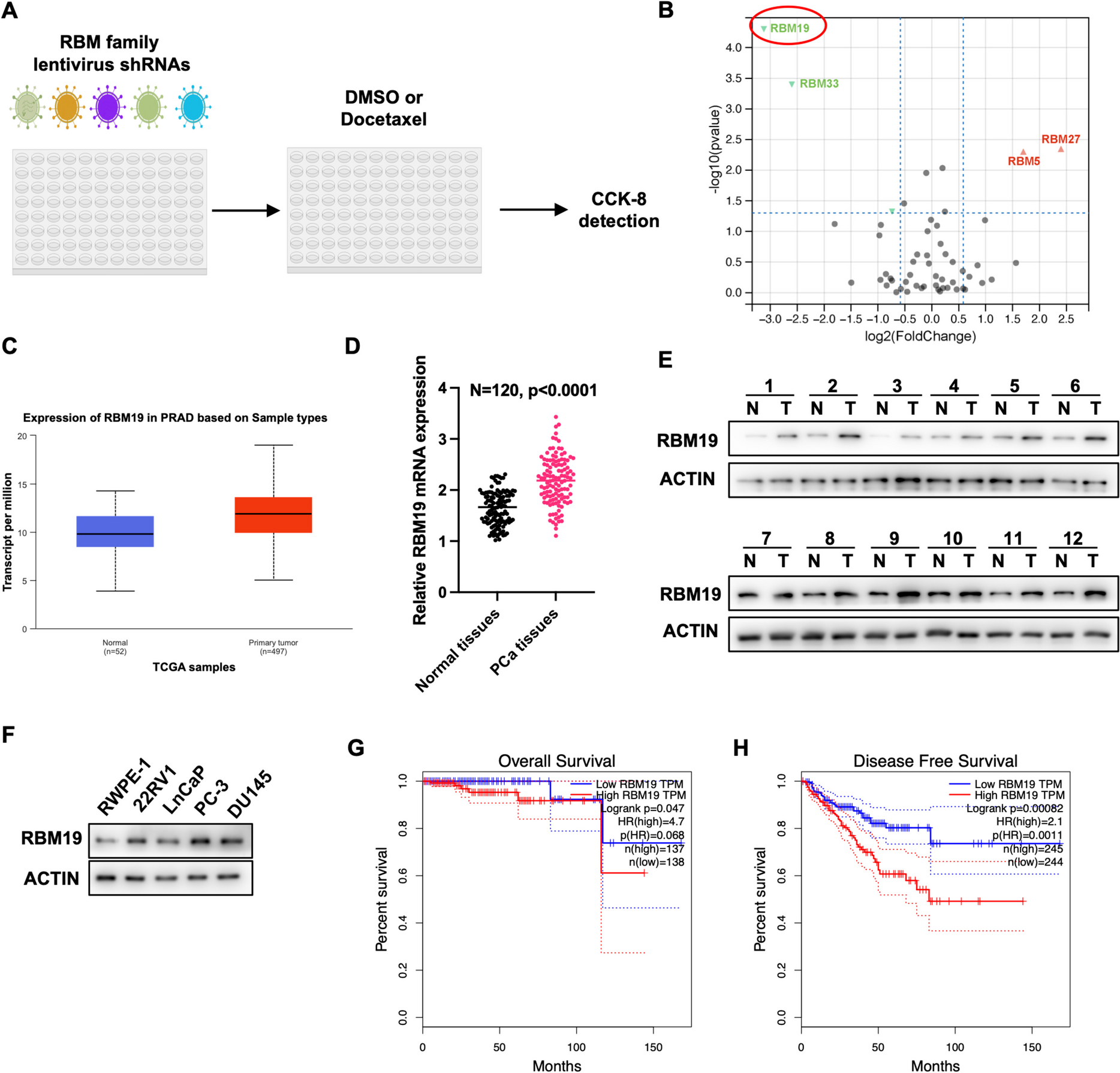



Previous studies have reported that members of RBM family involved in cancer progression (Cong et al. 2022; Li et al. 2021). Additionally, our group have revealed that RBM47 can restrain renal cell carcinoma progression (Su et al. 2023). To identified the specific RBM proteins involved in docetaxel treatment in PCa cells, an RNAi screen was applied in 22RV1 cells using an shRNA library targeting 54 members of the RBM family (Fig. 1A). Docetaxel or DMSO was applied in 22RV1 cells with specific RBM proteins depletion. After 4 days, the cell vitality of 22RV1 cells in response to docetaxel was detected using CCK-8 assays. The result showed that knockdown of RBM19 and RBM33 increased the sensitivity of 22RV1 to docetaxel, while silencing of RBM5 and RBM27 decreased the sensitivity of 22RV1 to docetaxel (Fig. 1B). Among these 4 RBM proteins, RBM19 was selected for further exploration due to its significant effect on 22RV1 cells under docetaxel treatment.
Fig. 1
RBM19 is linked to docetaxel regulation and is elevated in PCa tissues. A An RNAi screen was performed using shRNA library targeting 54 members of the RBM family to identify specific RBM proteins involved in docetaxel treatment in PCa cells. B Volcano plot indicated that silencing of RBM19 and RBM33 lead to an increased sensitivity of 22RV1 cells to docetaxel, while silencing of RBM5 and RBM27 result in a decrease in sensitivity. C TCGA-PRAD database analysis showing the high expression of RBM19 in PCa tissues. D qRT-PCR assay showing the elevated expression of RBM19 in PCa tissues compared with normal tissues. E Western blot assay showing the elevated expression of RBM19 in PCa tissues compared with normal tissues. F Western blot assay showing the elevated expression of RBM19 in PCa cell lines compared with the normal cell line, RWPE-1. G TCGA-PRAD database analysis showing the overall survival of PCa patients with RBM19 high-expression or low-expression. H TCGA-PRAD database analysis showing the disease-free survival of PCa patients with RBM19 high-expression or low-expression. Data were presented at mean ± S.D. from at least three independent experiments
Subsequently, we assessed the clinical significance of RBM19 in PCa patients. By evaluating the expression of RBM19 in TCGA-PRAD database, we found that RBM19 level was higher in PCa tissues than in normal tissues (Fig. 1C). Furthermore, RBM19 expression was positively associated with the patients’ Gleason score and nodal metastasis status (Figs. S1A-S1D). We then detected the expression of RBM19 in 120 paired PCa tissues using qRT-PCR, which revealed that RBM19 was up-regulated in PCa tissues compared with adjacent normal specimens (Fig. 1D). Moreover, the protein level of RBM19 was also elevated in 12 paired PCa tissues (Fig. 1E). We also assessed the expression of RBM19 in PCa cell lines and the prostate normal epithelial cell line, RWPE-1, and the result indicated that the elevated expression of RBM19 in PCa cell lines (Fig. 1F). Finally, the expression of RBM19 was found to be negatively correlated with the prognosis of PCa patients in TCGA-PRAD database (Figs. 1G and 1H). Together, these data show that RBM19 is linked to the docetaxel treatment in PCa cells and might act as an oncogene in prostate cancer.
RBM19 increased the cell vitality of PCa cells under docetaxel treatmentTo investigate RBM19’s role in PCa progression under docetaxel treatment, we first detected the expression RBM19 under docetaxel treatment. Surprisingly, we found that docetaxel treatment did not interfere the expression of RBM19 (Figs. S2A and S2B). Next, the RBM19 shRNAs and overexpression plasmids were constructed, and their silencing or overexpression efficiency were detected using western blot assays (Figs. S2C and S2D). Next, we employed the CCK-8 and colony formation assays to evaluate the biological role of RBM19 in prostate cancer cells under docetaxel treatment. The CCK-8 assays indicated that silencing of RBM19 significantly decreased the cell vitality of prostate cancer cells under docetaxel treatment (Fig. 2A), while RBM19 overexpression increased the cell vitality of prostate cancer cells under docetaxel treatment (Fig. 2B). Meanwhile, colony formation assays also revealed that RBM19 knockdown significantly decreased the proliferation ability of 22RV1 and PC-3 cells, whereas overexpression of RBM19 increased the proliferation ability of PCa cells (Figs. 2C and 2D). Additionally, cell cycle assays showed that silencing of RBM19 prevented the transition of PCa cells from G1 phase to S phase (Fig. 2E), while overexpression of RBM19 promoted this transition (Fig. 2F). Totally, these data reveal that RBM19 increases the vitality of PCa cells under docetaxel treatment.
Fig. 2
RBM19 increased the cell vitality of PCa cells under docetaxel treatment. A CCK-8 assay showing the cell vitality of PCa cells with RBM19 knockdown under docetaxel treatment. B CCK-8 assay showing the cell vitality of PCa cells with RBM19 overexpression under docetaxel treatment. C Colony formation assay showing the proliferation ability of PCa cells with RBM19 knockdown under docetaxel treatment. D Colony formation assay showing the proliferation ability of PCa cells with RBM19 overexpression under docetaxel treatment. E Cell cycle assay showing the cell cycle phase change with RBM19 knockdown under docetaxel treatment. F Cell cycle assay showing the cell cycle phase change with RBM19 overexpression under docetaxel treatment. Data were presented at mean ± S.D. from at least three independent experiments
RBM19 promoted the stability of LncRNA SNHG21Early research has indicated that RBM family proteins can modulate pre-mRNA splicing, affect mRNA stability, and interact with LncRNA (Qin et al. 2022; Su et al. 2023; Ye et al. 2018). This research tends to explore whether RBM19 bind to LncRNAs to influence prostate cancer progression. To identified the RBM19-regulated LncRNAs, we used specific RBM19 antibodies and negative control IgG antibodies to perform RNA Immunoprecipitation (RIP)-seq assays in both 22RV1 and PC-3 cell lines. Subsequently, we compared the RBM19 groups to the IgG control groups within each cell line to obtain the corresponding Log2FC values and p-values for each group. A total of 516 LncRNAs in 22RV1 cells and 581 LncRNAs in PC-3 cells were selected based the following criteria: (1) Log2FC > 1; (2) P < 0.05 (Fig. 3A). Among these RBM19-binding LncRNAs, 36 LncRNAs were found to interact with RBM19 in both 22RV1 and PC-3 cells. Among these 36 LncRNAs, we chose the top 11 LncRNAs with highest binding capacity for further study. Subsequently, RBM19-RIP-qPCR assay verified that SNHG21 exhibited the highest binding capacity with RBM19 (Fig. 3B). Furthermore, we performed the RNA pulldown assays using SNHG21 probe and its antisense probe. The result confirmed that SNHG21 could specifically bind to RBM19 in 22RV1 and PC-3 cells (Fig. 3C). To elucidate which domain of RBM19 contributes to the interaction between RBM19 and SNHG21, we construct several Flag-tagged truncations of RBM19 and performed Flag-RIP-qPCR assay. The result highlighted the RRM1-4 regions of RBM19 were critical for the interaction between RBM19 and SNHG21 (Fig. 3D). Consequently, we detected the role of docetaxel in the interaction between RBM19 and SNHG21. RBM19-RIP-qPCR assay indicated that RBM19 interacted with SNHG21, while docetaxel treatment enhanced their interaction (Fig. 3E). Moreover, we explored the effect of RBM19 and SNHG21 interaction. RT-qPCR assays revealed that RBM19 knockdown decreased the expression of SNHG21, while RBM19 overexpression increased the expression of SNHG21 (Fig. 3F). Considering that RBM19 could increase the expression of SNHG21, we, therefore, hypothesized that RBM19 could enhance the stability of SNHG21 to increased its expression. LncRNA stability assay was then performed to verified this hypothesis, showing that inhibition of RBM19 decreased the stability of LncRNA SNHG21 after actD treatment, while RBM19 increased the stability of LncRNA SNHG21 (Fig. 3G). Additionally, we found that docetaxel treatment increased SNHG21 expression in a dose-dependent manner (Fig. 3H), which is agreement with the previous finding that docetaxel treatment enhanced the interaction between RBM19 and SNHG21. Furthermore, qRT-PCR showed that SNHG21 was up-regulated in PCa tissues compared with adjacent normal specimens (Fig. 3I). Consequently, we identified that a positively correlation between RBM19 and SNHG21 expression of our 120 paired PCa tissues (Fig. 3J), as well as in the TCGA-PRAD database (Fig. 3K). Collectively, these data show that RBM19 binds to LncRNA SNHG21, and their interaction enhances SNHG21 stability.
Fig. 3
RBM19 promoted the stability of LncRNA SNHG21. A Venn diagram showing there were 36 common LncRNA both interact with RBM19 in RIP-seq data of 22RV1 and PC-3. B RIP-qPCR assays showing the binding capacity between LncRNAs and RBM19. C RNA pulldown assay showing the binding capacity between SNHG21 and RBM19. D Upper, the diagram illustrating the truncations of RBM19 proteins. Lower, RIP-qPCR assay showing the binding capacity between SNHG21 and Full length or truncations of RBM19. E RIP-qPCR showing the binding capacity between RBM19 and SNHG21 under docetaxel treatment. F RT-qPCR showing the expression of SNHG21 with RBM19 knockdown or overexpression. G LncRNA stability assay showing the stability of LncRNA in PCa cells with RBM19 knockdown or overexpression under actD treatment. H RT-qPCR showing the expression of SNHG21 in PCa cells under docetaxel treatment. I qRT-PCR assay showing the elevated expression of RBM19 in PCa tissues compared with normal tissues. J Correlation analysis showing the positive correlation between SNHG21 and RBM19 in 120 paired PCa tissues. K TCGA-PRAD database analysis showing the positively correlation between SNHG21 and RBM19. Data were presented at mean ± S.D. from at least three independent experiments. **p < 0.01, ***p < 0.001
SNHG21 increased the cell vitality of PCa cells under docetaxel treatmentWe further investigated the role of SNHG21 in PCa progression. First, by analyzing the expression of SNHG21 in TCGA-PRAD database, we found SNHG21 was up-regulated in PCa tissues (Fig. 4A). We then detected the expression of SNHG21 in our 120 paired PCa tissues, and the result showed that SNHG21 was up-regulated in PCa tissues compared with adjacent normal tissues (Fig. 4B). Furthermore, we observed that RBM19 expression was elevated in PCa cell lines compared with the prostate normal epithelial cell line, RWPE-1 (Fig. 4C). Also, elevated level of SNHG21 was linked to unfavorable outcomes in PCa patients in TCGA-PRAD database (Fig. 4D).
Fig. 4
SNHG21 increased the cell vitality of PCa cells under docetaxel treatment. A TCGA-PRAD database analysis showing the high expression of SNHG21 in PCa tissues. B qRT-PCR assay showing the elevated expression of SNHG21 in PCa tissues compared with normal tissues. C qRT-PCR assay showing the elevated expression of SNHG21 in PCa cell lines compared with normal cell line, RWPE-1. D TCGA-PRAD database analysis showing the disease-free survival of PCa patients with SNHG21 high-expression or low-expression. E CCK-8 assay showing the cell vitality of PCa cells with SNHG21 knockdown under docetaxel treatment. F CCK-8 assay showing the cell vitality of PCa cells with SNHG21 overexpression under docetaxel treatment. G Colony formation assay showing the proliferation ability of PCa cells with SNHG21 knockdown or overexpression under docetaxel treatment. H CCK-8 assay showing the cell vitality of RBM19 knockdown PCa cells with SNHG21 overexpression or not under docetaxel treatment. I Colony formation assay showing the proliferation ability of RBM19 knockdown PCa cells with SNHG21 overexpression or not under docetaxel treatment. Data were presented at mean ± S.D. from at least three independent experiments. **p < 0.01, ***p < 0.001
Next, we employed the CCK-8 and colony formation assays to evaluate the biological role of SNHG21 in PCa cells under docetaxel treatment. The CCK-8 assays indicated that knockdown of SNHG21 restrained the cell vitality of prostate cancer cells under docetaxel treatment (Fig. 4E), while overexpression of SNHG21 increased the cell vitality of PCa cells under docetaxel treatment (Fig. 4F). Moreover, similar results were observed in colony formation assays (Figs. 4G and S3A). Furthermore, we explored the role of SNHG21 in RBM19-regulated PCa progression under docetaxel treatment. CCK-8 assay indicated that RBM19 silencing decreased the cell vitality of 22RV1 and PC-3 cells, while overexpression of SNHG21 rescued this effect (Fig. 4H). Meanwhile, colony formation assay indicated that silencing of RBM19 inhibited the proliferation rate of PCa cells, while SNHG21 overexpression rescued this effect (Figs. 4I and S3B). In summary, the data indicate that SNHG21 enhances PCa cells proliferation under docetaxel condition, and RBM19 regulates PCa cells sensitivity to docetaxel depending on SNHG21.
SNHG21 interacted with PIM1 and prevent it from ubiquitin-protease dependent degradationRecently, numerous LncRNAs were reported to exert their function through binding with functional proteins (Barth et al. 2020; Peng et al. 2017). To identify the protein partners of SNHG21, we performed RNA pulldown assays followed by mass spectrometry. The result showed that 87 proteins specifically bind to LncRNA SNHG21 (Fig. 5A). Among these proteins, PIM1 exhibited the highest binding affinity to SNHG21. Moreover, RBM19 was also found in the mass spectrometry results. Subsequently, RNA pulldown assays and PIM1-RIP-qPCR assay verified the interaction between SNHG21 and PIM1 (Fig. 5B and 5C). We then explored the effect of PIM1 and SNHG21 interaction. RT-qPCR assays revealed that silencing of PIM1 did not change the expression of SNHG21 (Fig. 5D). However, western blot assay revealed that SNHG21 silencing decreased PIM1 expression, while SNHG21 overexpression increased PIM1 expression in PCa cells (Fig. 5E). Moreover, changing the expression of SNHG21 had minimal effect on PIM1 mRNA expression (Fig. S4A). Next, we tend to explore the specific mechanism by which SNHG21 regulated PIM1 protein expression. Initially, we treated PCa cells with cycloheximide (CHX), a protein translation inhibitor, to block PIM1 synthesis. Then, we detected PIM1 protein expression with SNHG21 knockdown or overexpression. The results indicated that SNHG21 knockdown decreased the protein half-time of PIM1 (Fig. 5F), while overexpression of SNHG21 increased the protein half-time of PIM1 (Fig. 5G). Furthermore, we treated PCa cells with proteasome inhibitor, MG132, or autophagy inhibitor, chloroquine, respectively to detected the protein expression of PIM1, and the result indicated that treated with MG132 increased the expression of PIM1 in PCa cells (Fig. 5H). These data suggest that SNHG21-induced PIM1 degradation might be dependent on the ubiquitin–proteasome pathway. Subsequently, we transfected PCa cells with ubiquitin and then detected the ubiquitination level of PIM1. The result revealed that SNHG1 silencing increased the ubiquitination level of PIM1, while SNHG21 overexpression decreased the ubiquitination level of PIM1 (Fig. 5I). Furthermore, we detected the PIM1 expression under docetaxel treatment, and the result demonstrated that docetaxel treatment elevated the PIM1 expression in a dose-dependent manner (Fig. 5J). Finally, we examined PIM1 protein expression in 12 paired PCa specimens. Our results showed that PIM1 was up-regulated in PCa tissues (Fig. 5K). Furthermore, a positive correlation was observed between PIM1 and RBM19 protein expression levels in PCa tissues, as determined by grayscale analysis (Fig. 5L). Altogether, these data reveal that SNHG21 interacts with PIM1 and prevents it from ubiquitin–proteasome-dependent degradation.
Fig. 5
SNHG21 interacted with PIM1 and prevent it from ubiquitin-protease dependent degradation. A The SNHG21-binding proteins identified by mass spectrometry were shown in the chart. B RNA pulldown assay showing the binding capacity between SNHG21 and PIM1. C RIP-qPCR showing the binding capacity between SNHG21 and PIM1. D RT-qPCR showing the expression of SNHG21 in PCa cell with PIM1 knockdown or overexpression. E Western blot assay showing the expression of PIM1 in PCa cells with SNHG21 knockdown or overexpression. F Western blot assay showing the protein stability of PIM1 in SNHG21 knockdown or not PCa cells treated with CHX (50 μg/mL). G Western blot assay showing the protein stability of PIM1 in SNHG21 overexpression or not PCa cells treated with CHX (50 μg/mL). H Western blot assay showing the expression of PIM1 in SNHG21 overexpression or knockdown PCa cells treated with MG132 (50 μg/mL) or CQ (20 μg/mL). I Flag-IP followed by western blot assay showing the ubiquitination levels of PIM1 protein in 22RV1 cells with SNHG21 overexpression or knockdown. J Western blot assay showing the expression of PIM1 in PCa cells treated with docetaxel. K Western blot assay showing the elevated expression of PIM1 in PCa tissues compared with normal tissues. L Correlation analysis showing the positive correlation between RBM19 and PIM1 in 12 paired PCa tissues. Data were presented at mean ± S.D. from at least three independent experiments. **p < 0.01, ***p < 0.001
SNHG21 promoted mitochondria homeostasis dependent on PIM1Previous studies have revealed that PIM1 kinase is a key factor that participates in cancer cells survival and proliferation in various types of cancers (Cao et al. 2019; Chauhan et al. 2020). Moreover, PIM1 is involved in the mitochondrial dynamics and ROS production (Chauhan et al. 2020; Song et al. 2018). Therefore, we hypothesized that PIM1 regulated mitochondrial homeostasis in prostate cancer. First, the PIM1 overexpression plasmids were constructed, and the overexpression efficiency were detected using western blot assays (Fig. S5A). Then, ROS levels were then detected using flow cytometry, and the results revealed that SNHG21 silencing increased the total ROS levels in PCa cells, while overexpression of PIM1 could rescue this promotion effect (Fig. 6A). Moreover, luminescence-based assays indicated that SNHG21 silencing decreased the NADPH and GSH levels in PCa cells (Figs. 6B and 6C). These data show that SNHG21 promotes mitochondrial homeostasis through PIM1.
Fig. 6
SNHG21 promoted mitochondria homeostasis dependent on PIM1. A Flow cytometry showing the ROS levels of SNHG21 knockdown PCa cells with PIM1 overexpression or not. B GSH was detected in SNHG21 knockdown PCa cells with PIM1 overexpression or not. C NADHP was detected in SNHG21 knockdown PCa cells with PIM1 overexpression or not. D CCK-8 assay showing the cell vitality of SNHG21 knockdown PCa cells with PIM1 overexpression or not under docetaxel treatment. E Colony formation assay showing the proliferation ability of SNHG21 knockdown PCa cells with PIM1 overexpression or not under docetaxel treatment. Data were presented at mean ± S.D. from at least three independent experiments. **p < 0.01, ***p < 0.001
Subsequently, a CCK-8 assay showed that SNHG21 knockdown decreased the cell vitality of PCa cells under docetaxel treatment, while overexpression of PIM1 could rescue this effect (Fig. 6D). Meanwhile, a colony formation assay indicated that silencing of SNHG21 decreased the proliferation ability of PCa cells under docetaxel treatment, while overexpression of PIM1 rescued this effect (Figs. 6E and S5B). These data indicate that SNHG21 regulates PCa cells sensitivity to docetaxel through PIM1.
Combination of RBM19 knockdown and docetaxel had a stronger tumor suppressor roleThe above data show that silencing of RBM19 can suppress the progression of PCa cells under docetaxel treatment in vitro. Thus, we investigated the role of RBM19 knockdown in vivo and whether the combination of RBM19 knockdown and docetaxel have a synergistic effect on the growth of PCa in vivo. The result showed that knockdown of RBM19 or docetaxel treatment significantly restrained 22RV1 cells proliferation in vivo; moreover, RBM19 knockdown together with docetaxel had a better therapeutic effect on tumor proliferation (Figs. 7A and 7C). Furthermore, we observed that low level of PIM1 in tumor tissues in the RBM19 knockdown group and both RBM19 knockdown and docetaxel treatment group (Fig. 7D). Moreover, the RBM19 knockdown and docetaxel treatment group exhibited the lowest Ki-67 levels (Fig. 7D). Totally, these data indicated that the combination of knockdown of RBM19 and docetaxel treatment had a synergistic inhibitory effect on the growth of PCa.
Fig. 7
Combination of RBM19 knockdown and docetaxel had a stronger tumor suppressor role. A Image of xenograft tumor in each group. B Tumor volume of xenograft tumor in each group. C Tumor weight of xenograft tumor in each group. D IHC staining showing the expression of RBM19, PIM1, and Ki-67 in each group. Data were presented at mean ± S.D. from at least three independent experiments
留言 (0)