
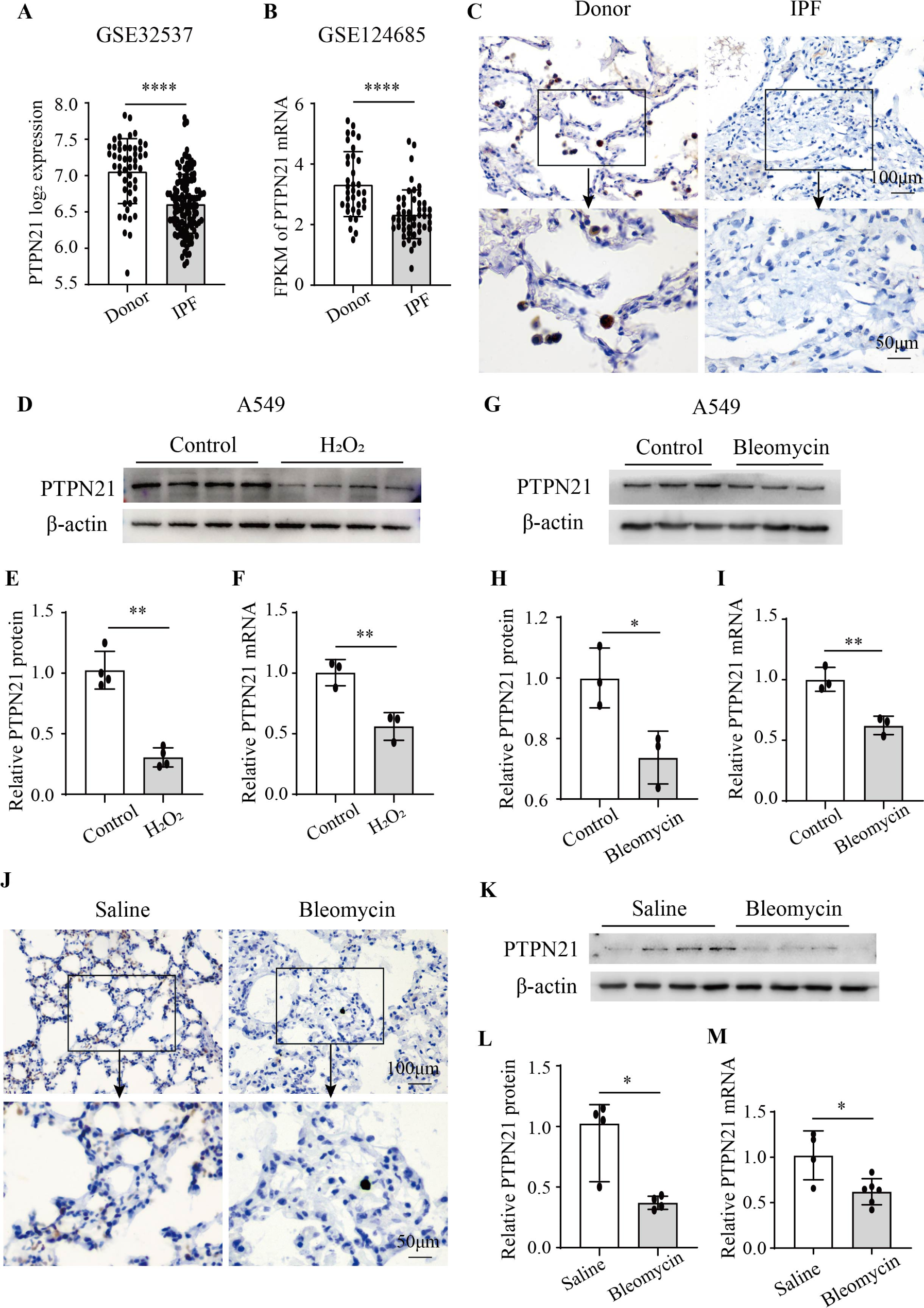
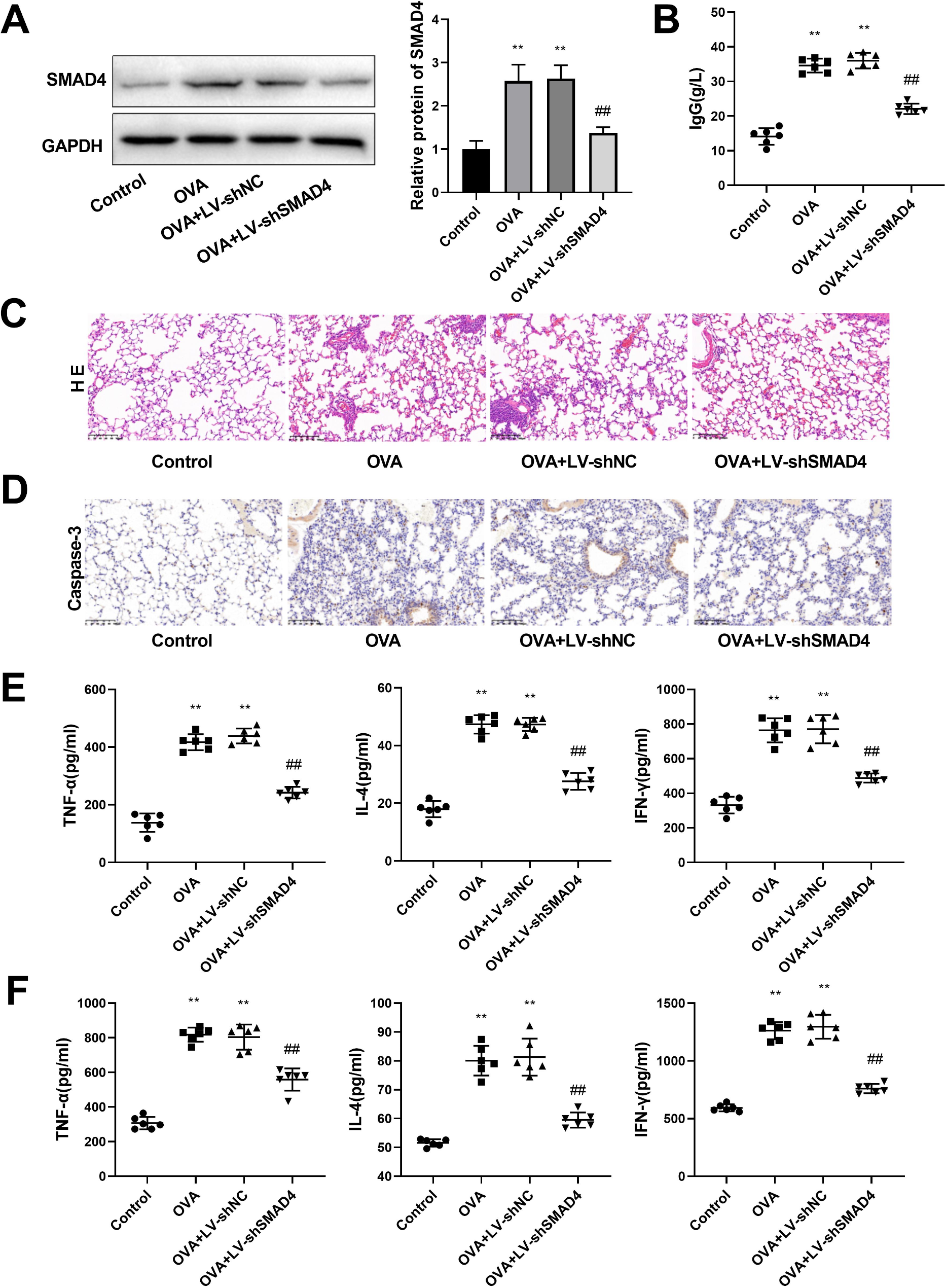
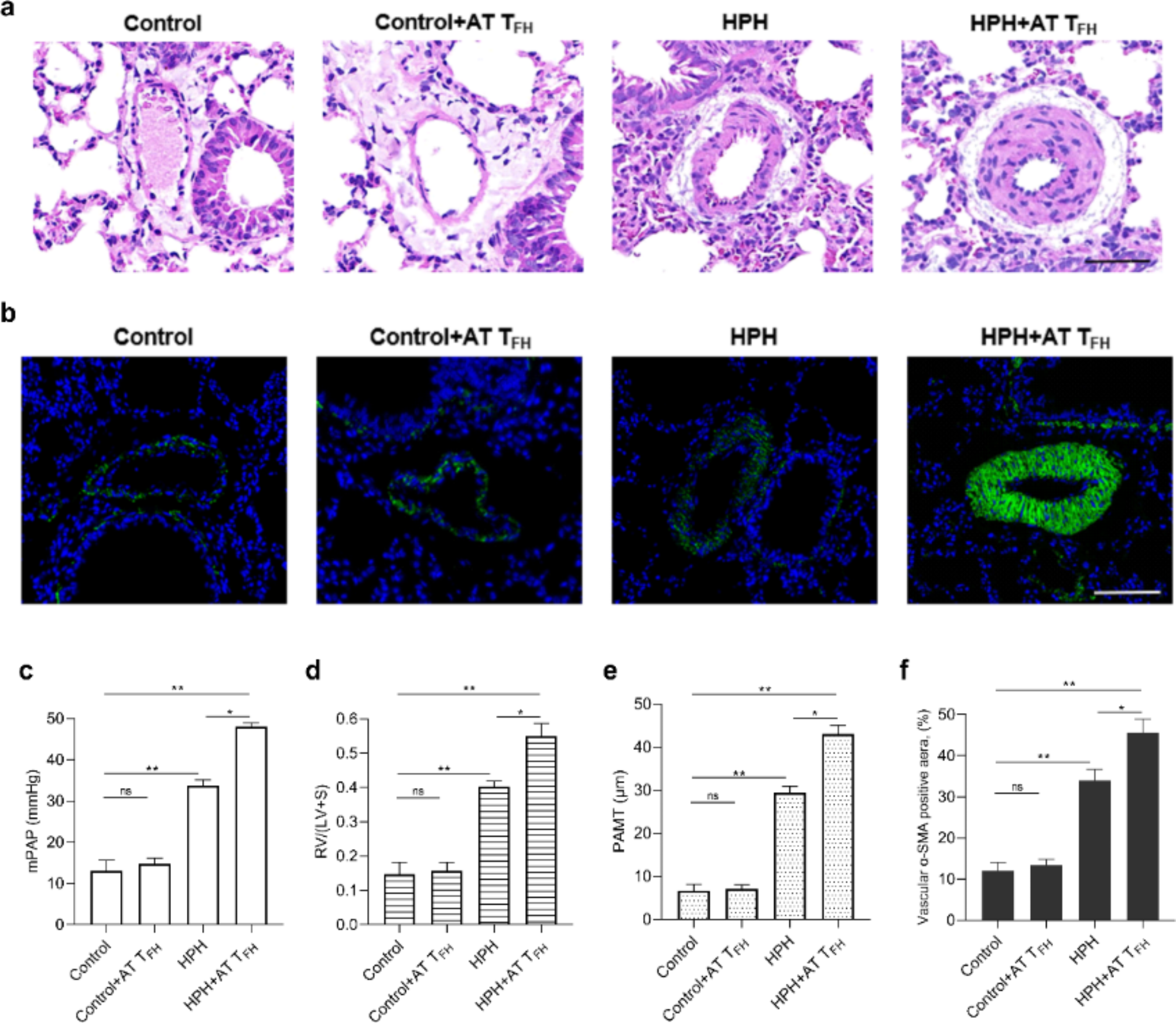
記住我
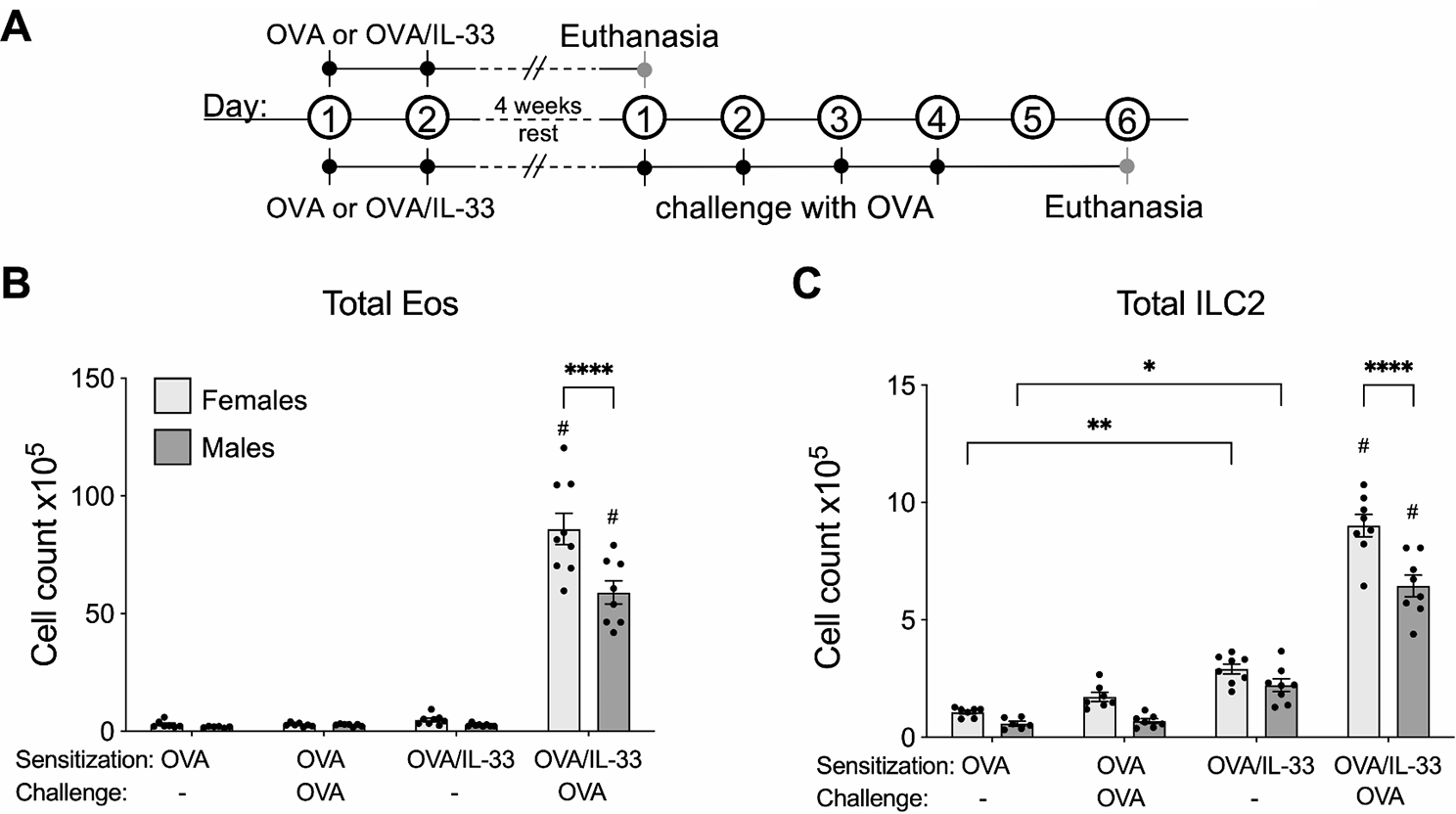
To better understand the influence of sex on the fate of IL-33-treated lung ILC2s in type 2 inflammation and allergic airways disease, we first examined ILC2 responses in a murine asthma model. Adult mice were treated with OVA alone (as the negative control) or OVA/IL-33 delivered to the lungs, after which mice were rested for 4 weeks and then either sacrificed directly or challenged with OVA to reactivate OVA-specific T cells prior to sacrifice (Fig. 1A). This model is similar to those of the Croft lab, showing that addition of lipopolysaccharide or a Nod2 agonist (muramyl dipeptide) to OVA at the time of antigen priming through the lung reduces induction of OVA-induced tolerance and, at the same time, promotes Th2 differentiation and type 2 inflammation following OVA challenge [24, 25]. Consistent with this, four weeks after initial priming, eosinophils were not elevated in mice sensitized with OVA alone, whether challenged with OVA or not. However, following OVA challenge of mice that had been sensitized with OVA/IL-33, eosinophils in both males and females were increased, to levels that were significantly greater in females (Fig. 1B) (exhibiting sex differences similar to those in house dust mite models of allergic airways disease) [14, 26].
Fig. 1
ILC2s persist in the lungs of male and female Balb/c mice for up to 4 weeks post OVA/IL-33 delivery. (A) Male and female Balb/c mice were sensitized with OVA or OVA/IL-33 once per day for 2 days. Mice were rested for 4 weeks and then either sacrificed directly or challenged with OVA once per day for each of 4 days and euthanized 48 h later, after which lungs were harvested for flow cytometry analysis. Absolute count of (B) eosinophils and (C) ILC2s in the lung. Data are the combination of 2 independent experiments, presented as mean ± SEM, with 6–8 mice/group. Outcomes were assessed by two-way ANOVA, followed by Tukey’s post hoc test. *p < 0.05, **p < 0.01, ****p < 0.0001. #: significantly different than all other groups within the same sex
Mice treated with OVA alone had negligible differences in total ILC2s, whether they were from males or females, and whether they were examined before or after OVA challenge (Fig. 1C). However, even without OVA challenge, ILC2s in mice sensitized with OVA/IL-33 were significantly increased, to levels that were similar in males and females (Fig. 1C). Following OVA challenge of OVA/IL-33 sensitized mice, ILC2s increased about 3-fold in both males and females, and similar to eosinophils, there were significantly more ILC2s in females compared to males (Fig. 1C). These data suggest that ILC2s persist in the lungs of both male and female mice for up to 4 weeks post activation.
IL-33-experienced ILC2s exhibit phenotypic differences in male and female miceSex-specific phenotypic differences in ILC2s in murine models have been noted both before and after activation. One notable difference is that in adult female mice there are larger numbers of ILC2s lacking expression of KLRG1 [13], findings we have confirmed [15]. Thus, we examined ILC2 populations expressing KLRG1 (or not) 4 weeks following exposure to OVA or OVA/IL-33. No sex differences were observed in the numbers of KLRG1+ ILC2s in any group, though the KLRG1+ population remained significantly elevated 4 weeks post OVA/IL-33 treatment in both males and females and increased further, and to the same extent, in mice of each sex upon OVA challenge (Fig. 2A). Moreover, compared to males, KLRG1– ILC2s were elevated in females, though a statistically significant sex difference was present only after OVA challenge of OVA/IL-33 sensitized mice. In addition, unlike ILC2s expressing KLRG1, the KLRG1– ILC2 population was not significantly elevated 4 weeks after OVA/IL-33 exposure (in mice of either sex) (Fig. 2B).
Fig. 2
IL-33-experienced ILC2s exhibit phenotypic differences in male and female mice. Mice were treated as described in Fig. 1A, after which lungs were harvested for flow cytometry analysis. Absolute count of (A) KLRG1+ ILC2s (B) KLRG1– ILC2s (C) IL25R+KLRG1+ ILC2s (D) IL-25R+KLRG1– ILC2s in the lung. MFI of ST2 on (E) KLRG1+ ILC2s (F) KLRG1– ILC2s. Data are the combination of 2 independent experiments, presented as mean ± SEM, with 6–8 mice/group for A-D and representative of 2 independent experiments with 6–8 mice/group in E and F. Outcomes were assessed by two-way ANOVA, followed by Tukey’s post hoc test. *p < 0.05, **p < 0.01, ****p < 0.0001. #: significantly different than all other groups within the same sex
Because upregulation of IL-25R has previously been associated with memory-like responses in ILC2s [19], we examined whether levels of this receptor were increased in IL-33-experienced ILC2s before or after OVA challenge, using the gating strategy presented in Fig S1. Following OVA challenge of OVA/IL-33-sensitized mice, the total number of IL-25R-positive ILC2s, expressing KLRG1 or not, was significantly increased only in female mice (Fig. 2C and D). Interestingly, there was also a clear trend for these cells to increase 4 weeks after OVA/IL-33 administration and prior to OVA challenge, again selectively in females. No change in the median fluorescence intensity (MFI) of IL-25R was detected (data not shown). Finally, while we defined ILC2s based on their ST2 expression (Fig S1), we also found that, 4 weeks after OVA/IL-33 exposure, ILC2s in female mice expressed higher levels of ST2; an increase that was present in both KLRG1+ and KLRG1– ILC2s (Fig. 2E and F), but absent in their male counterparts. This increase in ST2 expression in ILC2s from female mice was no longer apparent following OVA challenge. These data suggest that, one month after initial activation, IL-33-experienced ILC2s in female mice are poised to respond in a more robust manner to secondary challenge with IL-33 or IL-25.
Sex differences in cytokine producing ILC2s are due to KLRG1– ILC2s in female miceWe also quantified cytokine producing ILC2s in mice exposed to OVA or OVA/IL-33, with and without OVA challenge. Cytokine producing ILC2s were not significantly elevated 4 weeks after OVA/IL-33 administration in either males or females; however, OVA challenge induced a significant increase in the number of IL-13+ KLRG1+ ILC2s in both males and females (Fig. 3A), but a similar increase in IL-13+ KLRG1– ILC2s was present only in females (Fig. 3C). IL-5+ KLRG1+ ILC2s tended to increase 4 weeks after OVA/IL-33 delivery, but OVA challenge did not further increase their number above that present in OVA/IL-33 treated mice (with no OVA challenge) (Fig. 3B). Females had significantly greater numbers of IL-5+ KLRG1– ILC2s compared to males in mice sensitized and challenged with OVA, as well as in mice exposed to OVA/IL-33, both before and after OVA challenge (Fig. 3D). While there was no significant increase in IL13+IL-5+KLRG1+ 4 weeks post OVA/IL-33 exposure in males or females, OVA challenge increased their number significantly in both sexes (Fig S2A). In addition, IL13+IL-5+KLRG1– ILC2s were elevated four weeks post OVA/IL-33 treatment in female mice only (Fig S2B) and these numbers increased significantly upon OVA challenge only in females (Fig S2B). Together, these data suggest that production of IL-13 and IL-5 is differentially regulated in ILC2s, with IL-13-producing ILC2s being much more responsive to OVA challenge in this model of Th2 adaptive immunity in the lung. Nevertheless, the larger number of IL-5+ KLRG1– ILC2s in females prior to OVA challenge was not sufficient to increase eosinophil numbers, as reflected in the small number of eosinophils present in all mice prior to OVA challenge (Fig. 1B). The larger number of cytokine-producing KLRG1– ILC2s in females likely contributes to the greater inflammatory responses in females compared to males, post OVA challenge.
Fig. 3
Cytokine producing KLRG1– ILC2s are greater in female mice. Mice were treated as described in Fig. 1A, after which lungs were harvested for flow cytometry analysis. Absolute count of (A) IL-13+KLRG1+ ILC2s (B) IL-5+KLRG1+ ILC2s (C) IL-13+KLRG1– ILC2s (D) IL-5+KLRG1– ILC2s in the lung. Data are the combination of 2 independent experiments, presented as mean ± SEM, with 6–8 mice/group. Outcomes were assessed by two-way ANOVA, followed by Tukey’s post hoc test. *p < 0.05, ***p < 0.001, ****p < 0.0001. #: significantly different than all other groups within the same sex
IL-33-experienced ILC2s are more responsive to IL-25 in female miceWe next examined the functional significance of IL-25R expression on IL-33-experienced ILC2s. Mice were treated with OVA or OVA/IL-33 as above. Four weeks later, OVA-sensitized mice were challenged with a single low dose of IL-25 and OVA/IL-33 sensitized mice were challenged with IL-25 or control saline as shown in Fig. 4A. These treatment groups are designated OVA→IL-25, OVA/IL-33→saline, and OVA/IL-33→IL-25, respectively. Compared to OVA→IL-25 treated mice, both eosinophils and ILC2s were significantly increased in OVA/IL-33→IL-25 treated male and female mice (Fig. 4B, C). In addition, in OVA/IL-33→saline exposed mice, eosinophil numbers were not elevated (Fig. 4B), even while ILC2s were increased (though the increase was not statistically significant) (Fig. 4C), similar to our findings presented in Fig. 2A. Eosinophils and ILC2s were both significantly increased, selectively in female mice, treated with OVA/IL-33→IL-25 (Fig. 4B, C), whereas these cell populations did not increase in male mice. Similar changes were noted in the KLRG1+ ILC2 population (Fig. 4D) as well as the KLRG1– ILC2 population (Fig. 4E), though, as expected, larger numbers of KLRG1– ILC2s were present in females compared to males. Taken together, these data demonstrate that IL-33-experienced ILC2s in female mice are markedly responsive to a single challenge with IL-25, whereas their male counterparts were largely unaffected.
Fig. 4
IL-33-experienced ILC2s in female mice are more responsive to IL-25. (A) Male and female Balb/c mice were sensitized with OVA or OVA/IL-33 once per day for 2 days. Mice were rested for 4 weeks, and then OVA-treated mice were challenged once with IL-25, while OVA/IL33-treated mice were challenged once with saline or IL-25 before being euthanized 48 h later and harvesting lungs for flow cytometry analysis. Absolute count of (B) eosinophils (C) ILC2s (D) KLRG1+ ILC2s (E) KLRG1– ILC2s in the lung. Data are the combination of 2 independent experiments, presented as mean ± SEM, with 8–10 mice/group. Outcomes were assessed by two-way ANOVA, followed by Tukey’s post hoc test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. #: significantly different than all other groups within the same sex
We also examined whether IL-25 promoted cytokine production by OVA/IL-33-experienced ILC2s. No significant differences in numbers of cytokine producing ILC2s (whether they expressed KLRG1 or not) were present in OVA/IL-33→saline exposed mice (Fig. 5A-D). IL-13+KLRG1+ and IL-13+KLRG1– ILC2s were significantly increased in OVA/IL-33→IL-25 exposed females (Fig. 5A, C), whereas similar changes were absent in male mice. Interestingly, no significant differences were observed between males and females in the numbers of IL-5+KLRG1+ ILC2s in OVA/IL-33→IL-25 exposed mice, where increases were noted in both males and females (Fig. 5B). In addition, IL5+KLRG1– ILC2s were significantly increased only in OVA/IL-33→IL-25 treated female mice (Fig. 5D). Similar responses in ILC2 cytokine production were present in mice challenged with IL-33 (Fig S3A-D), providing evidence that enhanced responsiveness in females was not limited to IL-25.
Fig. 5
IL-25 increases cytokine production and number of IL-25R expressing ILC2s selectively in female mice. Mice were treated as described in Fig. 4A, after which lungs were harvested for flow cytometry analysis. Absolute count of (A) IL-13+KLRG1+ ILC2s (B) IL-5+KLRG1+ ILC2s (C) IL-13+KLRG1– ILC2s (D) IL-5+KLRG1– ILC2s (E) IL-25R+KLRG1+ ILC2s (F) IL-25R+KLRG1– ILC2s in the lung. Data are the combination of 2 independent experiments, presented as mean ± SEM, with 8–10 mice/group. Outcomes were assessed by two-way ANOVA, followed by Tukey’s post hoc test. **p < 0.01, ****p < 0.0001. #: significantly different than all other groups within the same sex
Our data presented in Fig. 2C showed a trend for an increase in the number of IL-25R+ ILC2s selectively in females 4 weeks post OVA/IL-33 administration. Thus, we hypothesized that IL-25 challenge would induce the expansion of IL-25R+ ILC2s in females. Our data show that a single challenge with IL-25 significantly increased the numbers of IL-25R+KLRG1+ and IL-25R+KLRG– ILC2s, as expected, only in female mice (Fig. 5E, F). IL25R+ ILC2s were also increased in response to secondary challenge with IL-33 (Fig S3E, F). Altogether, these data provide further evidence that IL-33-experienced ILC2s in female mice are much more responsive to secondary activation compared to ILC2s in males.
The expression of IL-25R on IL-33-experienced ILC2s is associated with greater propensity to produce type 2 cytokinesWe also examined how expression of IL-25R impacted responsiveness to secondary exposure to IL-25 or IL-33. While larger numbers of ILC2s lacking expression of IL-25R were present in both males and females (data not shown), ILC2s expressing IL-25R had a greater propensity to produce IL-13 and/or IL-5: compare black portion of circles in Fig. 6A and B for males and Fig. 6C and D for females. Moreover, the proportion of cytokine producing ILC2s (whether IL-25R+ or not) was greater in females compared to males: compare Fig. 6C to Fig. 6A and Fig. 6D to Fig. 6B. Similar results were observed in mice challenged with IL-33 (data not shown). Together, our data provide evidence that expression of IL-25R marks ILC2s that are poised to respond in an enhanced manner upon secondary activation, whether by IL-25 or IL-33.
Fig. 6
The expression of IL-25R on IL-33-experienced ILC2 is associated with a greater propensity for cytokine production in response to IL-25 challenge. Mice were sensitized with OVA/IL-33 and challenged with IL-25 as described in Fig. 4A, after which lungs were harvested for flow cytometry analysis. The frequency of ILC2s producing IL-13 and/or IL-5 (black portion of rings) or no cytokines (grey portion of rings) in (A) IL-25R+ ILC2s in male mice (B) IL-25R– ILC2s in male mice (C) IL-25R+ ILC2s in female mice (D) IL-25R– ILC2s in female mice. Data are the combination of 2 independent experiments, with 8–10 mice/group
Enhanced responsiveness of IL-33-experienced ILC2s to IL-25 is dependent upon STAT6STAT6 has recently been linked to memory responses in ILC2s [20]. Thus, we also examined whether the absence of STAT6 had an impact on the enhanced responsiveness to IL-25 of ILC2s previously exposed to IL-33. Wild-type and STAT6 KO female mice were treated with IL-33 and challenged, one month later, as above with control saline or IL-25. Similar to our findings presented in Figs. 4 and 5, our data show that IL-33-experienced KLRG1+ ILC2s from wild-type mice were significantly elevated 4 weeks after IL-33 exposure and further increased upon secondary activation with IL-25 (Fig. 7A, C, E; left panels). Increases were present in total ILC2s (Fig. 7A) as well as those producing IL-13 (Fig. 7C) or IL-5 (Fig. 7E). Similar increases were noted for KLRG1– ILC2s (Fig. 7B, D, F; left panels). In STAT6 KO mice, KLRG1+ ILC2s producing IL-13 or IL-5 were significantly increased 4 weeks after IL-33 exposure and a similar trend was observed for total KLRG1+ ILC2s (Fig. 7A, C, E; right panels), providing evidence that ILC2s from STAT6 KO mice retain the ability to respond to IL-33. However, delivery of IL-25 did not further enhance total or cytokine-producing KLRG1+ ILC2s in STAT6 KO mice (Fig. 7A, C, E; right panels). KLRG1– ILC2s from STAT6 KO mice were less responsive than their counterparts expressing KLRG1 (Fig. 7B, D, F; right panels). Altogether, these data suggest that the absence of STAT6 does not limit the ability of ILC2s to respond to IL-33, though it is required for enhanced responsiveness to IL-25, a response associated with memory like activity in IL-33-experienced ILC2s.
Fig. 7
Enhanced responsiveness of IL-33-experienced ILC2s to IL-25 is dependent upon STAT6. Mice were treated as described in Fig. 4A, replacing OVA with saline as described in methods, after which lungs were harvested for flow cytometry analysis. Absolute count of (A) KLRG1+ ILC2s (B) KLRG1– ILC2s (C) IL-13+KLRG1+ ILC2s (D) IL-13+KLRG1– ILC2s (E) IL-5+KLRG1+ ILC2s (F) IL-5+KLRG1– ILC2s in the lung. In each graph wild-type mice are on the left, and STAT6 KO mice are on the right. Data are the combination of 3 independent experiments in wild-type mice and, in STAT6 KO mice, 1 experiment for Sal-IL-25 group and IL-33-Sal group and the combination of 2 independent experiments for the IL-33-IL-25 group. Data are presented as mean ± SEM with 3–13 mice/group. Outcomes were assessed by one-way ANOVA, followed by Tukey’s post hoc test for mice of each genotype. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
留言 (0)