
記住我
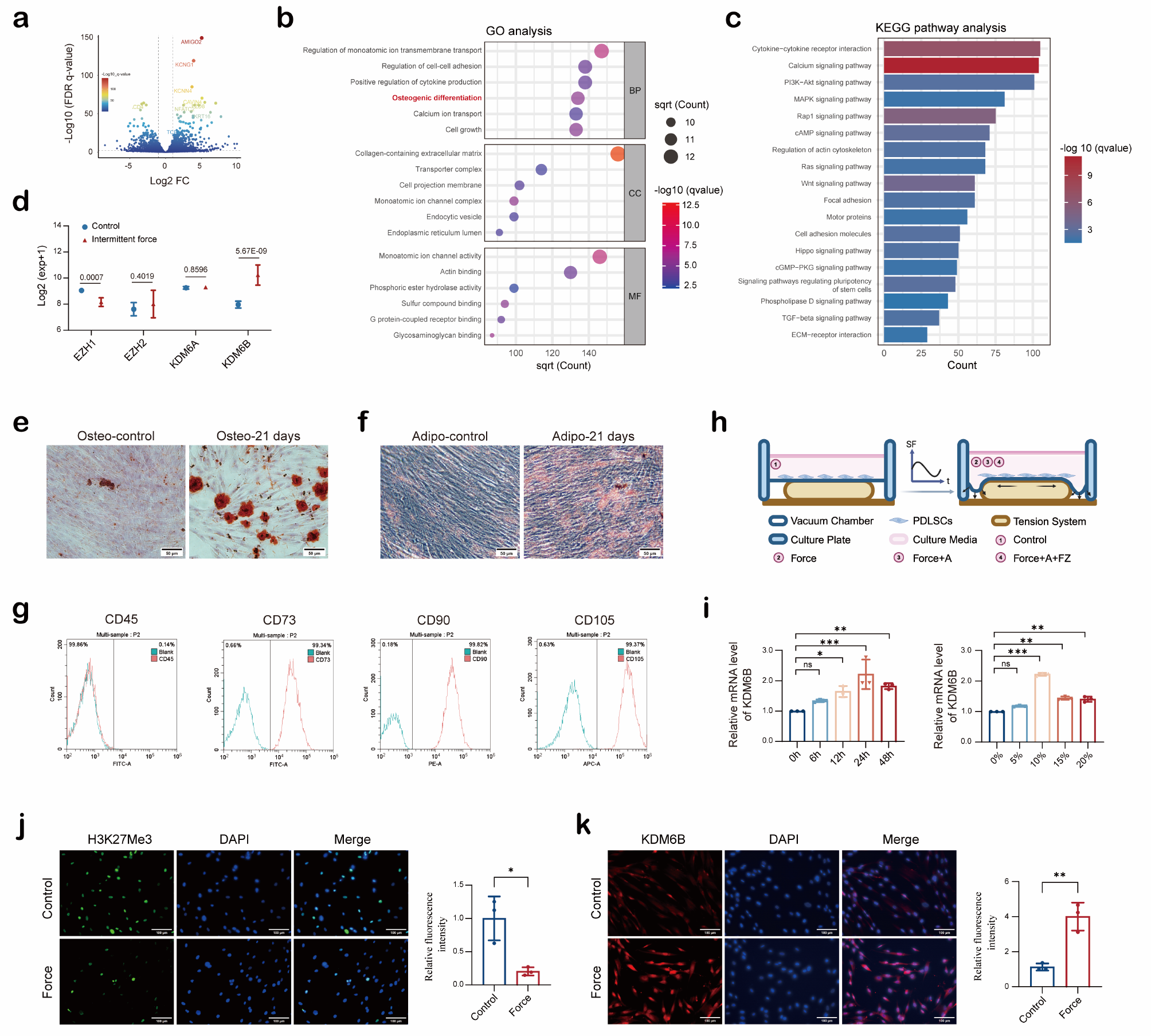





We first determined the in vivo persistence of MSCs delivered via SC injection compared to other injection routes. Bioluminescence imaging (BLI) was performed on healthy mice following IV, IP or SC injection of firefly luciferase-expressing MSCs (fluc-GFP+ MSC). The transduction efficiency of MSCs was quantified by flow cytometry indicated by the proportion of GFP-expressing cells (Fig. 1A, ~ 72%). The total number of cells to be injected was adjusted accordingly to ensure each mouse received 1 × 106 fluc-GFP+ MSCs per injection.
Fig. 1
SC injected MSCs display a longer dwell time at the site of injection. (A), MSCs were transduced with a firefly luciferase (fluc) vector that also encodes eGFP (fluc-GFP) to enable the quantification of transduction efficiency (indicated by %flu-GFP+) by flow cytometry. The cell suspension prepared for injection was adjusted to ensure 1 × 106 fluc-GFP+ MSCs per injection. (B) Representative bioluminescent images of luciferase-expressing MSCs after IV, IP and SC injections, with bar graphs present the changes in the radiance (n = 3 mice per group). (C) In vivo bioluminescent signal titration: various number of luciferase-expressing MSCs were injected via SC route into the hock to examine the lower threshold of the detection range. Data expressed as mean ± SEM; n = 3 mice per group
MSCs administered via the IV route undergo apoptosis in the lung, a process shown to be necessary for inducing anti-inflammatory effects [8]. As expected, IV-injected MSCs were detected in the lung immediately after injection, and a rapid decay of their bioluminescent signals was observed within 1 day, with complete clearance on day 3 (Fig. 1B). IP-injected MSCs remained in the peritoneum around the injection site and the signals persisted for a slightly longer period of time than IV-injected MSCs, with complete clearance on day 4 (Fig. 1B). Compared to the IV or IP injection groups, MSCs administered via the SC route into the hock displayed the longest in vivo dwell time, with complete clearance on day 5 (Fig. 1B). SC-injected MSCs remained locally at the injection site (hock), without evidence of migration away from the injection site.
A titration of SC fluc-GFP+ MSC injection dose demonstrated the lower threshold for the BLI detection range being 10,000 fluc-GFP+ MSCs (Fig. 1C). Hence, the gradual disappearance of BLI signals at the SC injection site may indicate clearance of exogenous MSCs at the injection site after 5 days, or a potential dissemination of less than 10,000 cells to elsewhere in the body which could not be effectively detected.
SC MSC injection into inflamed skin expands immunoregulatory IL-10+ macrophages in the draining LNIn inflammation and other disease states, immune responses are initiated and regulated in secondary lymphoid organs such as LNs. Studies characterizing the LN response to an acute inflammatory stimulus have commonly examined the popliteal lymph node draining the footpad where the inflammatory stimulus was injected [22]. The popliteal lymph nodes in mice are very small but easily located and separated from the skin and fascia [23]. Therefore, having confirmed that SC-injected MSCs remained at the injection site over a longer duration than IV- or IP-injected MSCs, we next established a localised acute inflammation model to provide an inflammatory microenvironment for SC-injected MSCs that would enable us to analyse the draining popliteal LN response.
TLR4 signalling pathway has been shown to be involved in establishing inflammatory hyperalgesia via the induction of pro-inflammatory cytokines such as TNF and IL-1β [24]. In a previous study, 100 ng of LPS (a TLR4 agonist) induced pro-inflammatory cytokines in the paw skin and inflammatory pain in the paw [24]. Due to institutional ethical constraints around animal welfare, we injected LPS into the hock, which is a non-weight bearing structure that drains to the same lymph node as the footpad, as a humane alternative to footpad injections [25]. We found 30 ng LPS injected into the hock was sufficient in inducing an acute inflammatory response in the skin, indicated by TNF, IL-1β and MCP-1 production, which peaked at 4 h post-LPS injection and resolved by 24 h (Fig. 2A), with concurrent expansion in total cellularity of the draining popliteal LN, increases in neutrophil and monocyte cell numbers from 4 h to 24 h (Fig. 2B) and elevation in their pro-inflammatory gene expression (TNF, IL-1β, IL-6 and MCP-1) (Fig. 2C). Surprisingly, a higher dose of LPS (100 ng) had a lower effect on cytokine production (Fig. 2A), possibly because higher doses of LPS induced a quicker inflammatory response that peaked at an earlier timepoint that was not captured here [24].
Fig. 2
Establishing an acute inflammation model with low-dose LPS. (A) C57BL/6 mice were SC-injected with 30 ng or 100 ng LPS in 50 µl PBS in the left hock. At 0, 4, 8, 24 h post-LPS injection, skin tissue around the LPS injection site was excised and homogenised for cytokine analysis. Changes in pro-inflammatory TNF, IL-1β and MCP-1 in the skin tissues over the time course. n = 3 mice per timepoint from one experiment. (B) Draining LNs were harvested from mice at 0, 4, 8, 24 h after SC injection of 30 ng LPS in 50 µl PBS in the left hock. Total LN, neutrophil and monocyte cellularity at various timepoints post-LPS injection. (C) Inflammatory gene expression in whole LN cell lysate and sorted LN neutrophils and monocytes at 24 h post-LPS injection. Fold-change differences in mRNA expression of inflammatory genes were plotted after normalising mRNA expression level to housekeeping genes (GAPDH and β-actin). Data expressed as mean ± SEM; UNT (untreated LN) n = 2–3 mice per timepoint; LPS (LPS dLN) n = 3–4 mice per timepoint
Adipose tissue-derived MSCs were then delivered via SC injection into the inflamed tissue for in vivo priming with TNF, IL-1β and MCP-1 (which are cross-reactive between mouse and human cells [26]) and the popliteal LN analysed at 24 h (Fig. 3A, B) when the SC-injected MSCs were still detectable at the site of injection (Fig. 1B). Although neutrophils and monocytes in the LN expanded in number in response to LPS injection and were the main producers of pro-inflammatory cytokines in this model (Fig. 2C), SC MSC injection did not affect the cellularity of these populations, nor the number of DCs or T cells (Fig. 3C). There was, however, a significant increase in B cells in the LN following SC injection of MSCs into the inflamed tissue (Fig. 3C, white bar), which was not found in the non-draining popliteal LN (LN from the non-inflamed hock; Fig. 3C, shaded bar). Regulatory B cells (Bregs) have been reported to attenuate inflammation and are characterised by their ability to produce IL-10 27, 28, 29, 30. We examined IL-10 production in B cells and found no increase in IL-10+ B cells (Fig. 3C), suggesting that SC MSC injection expanded the bulk B cell population rather than a specific regulatory B cell subtype.
Fig. 3
SC MSC injection expands IL-10-producing MerTK+ macrophages in the LN. (A) C57BL/6 mice were SC-injected with 30 ng LPS in 50 µl PBS in the left hock, and 4 h later 1 × 106 human adipose MSCs in the same hock. Draining (dLN; white bar) and non-draining LNs (ndLN; shaded bar) were harvested 20 h after MSC injection for analysis. (B) Representative flow cytometry plots showing the gating strategies for delineating myeloid and lymphoid populations in the mouse LNs. (C) Changes in total cellularity and cell number of different immune populations in LNs following SC MSC injection. Data expressed as mean ± SEM; n = 4–5 mice per group, representative of 2 independent experiments. One-way ANOVA, Tukey’s multiple comparison, comparing UNT, LPS and MSC dLNs; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. (D) Changes in cell number of different LN-resident macrophage subpopulations following MSC treatment. (E) Representative flow cytometry plots showing the identification of IL-10+ populations in LN macrophage subpopulations. (F) Changes in IL-10+ populations in LN macrophage subpopulations after LPS and SC MSC injection. Data expressed as mean ± SEM; n = 5 mice per group, representative of 2 independent experiments. One-way ANOVA, Tukey’s multiple comparison, comparing UNT, LPS and MSC dLNs; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant
In the lymph node, resident macrophages play crucial roles in immune regulation. Subcapsular sinus macrophages (SSM) act as gatekeepers by capturing foreign antigens in the lymph, whilst medullary sinus macrophages (MSM), medullary cord macrophages (MCM), tingible body macrophages and T cell zone macrophages (TZM) actively clear apoptotic cell debris, contributing to tissue homeostasis [19]. Following SC MSC injection, there was also an increase in MerTK+ SSM and TZM, but not F4/80+ MSM and MCM (Fig. 3D, white bar). None of these cellularity changes in response to LPS and MSC injection was observed in the non-draining popliteal LN (Fig. 3D, shaded bar).
LN-resident macrophages possess immunoregulatory function that protect against pathogenic infection and inflammatory insults [31, 32], and MerTk signalling in macrophages has been linked to anti-inflammatory outcomes [33, 34]. IL-10 has also been shown to shift macrophages toward an immunoregulatory and tolerogenic phenotype [35]. We therefore investigated whether the expansion of MerTK+ LN macrophages after SC MSC injection is associated with anti-inflammatory IL-10 production (Fig. 3E). The number of IL-10-producing macrophages remained unchanged in LPS mice, but SC MSC injection increased the number of IL-10-producing SSM and TZM in the draining popliteal LN (Fig. 3F, white bar). This increase was not observed in F4/80+ macrophages (Fig. 3F). IL-10+ macrophages also remained unchanged in the non-draining LN (Fig. 3F, shaded bar). Thus, SC MSC injection induced an increase in IL-10-producing MerTK+ macrophages but not F4/80+ macrophages in the draining LN.
SC MSC injection into inflamed skin expands activated Tregs with decreased PD-1 expression in the draining LNFollowing IV administration of MSCs, MSC-primed monocytes/macrophages have been shown to enhance the differentiation of T cells to Tregs [7, 36, 37], which have crucial immunoregulatory function in the LN [38,39,40]. The increase in IL-10+ MerTK+ macrophages in the LN after SC MSC injection prompted us to examine the immunomodulatory capacity of macrophages following SC MSC injection. MerTK+ macrophages (SSM and TZM), neutrophils, monocytes and DCs were isolated from the LN of MSC-treated mice and co-cultured with CTV-labelled CD4+CD25− T cells that were activated with anti-CD3/anti-CD28 (Fig. 4A). Activated T cells underwent proliferation, as indicated by CTV dilution (Fig. 4B). Neither macrophages, neutrophils, monocytes nor DCs suppressed T cell proliferation (Fig. 4C). However, analysis of Foxp3+ Tregs in the co-culture (Fig. 4D) revealed that macrophages from the LN of SC MSC-injected mice increased the proportion of Tregs, compared to macrophages from untreated mice (Fig. 4E). The expansion of Tregs was not observed with neutrophils, monocytes or DCs from the same LN of SC MSC-injected mice (Fig. 4E). Thus, MerTK+ macrophages from the LN of mice that received SC MSC injection exhibited an enhanced capacity to differentiate and/or expand Tregs.
Fig. 4
SC MSC injection expands activated Tregs in the LN. (A) CD45+Ly6G+SSC-Ahi neutrophils, Ly6Clo/hiCD11b+ monocytes, CD11c+MHCII+ DCs and CD11b+MerTk+ macrophages were purified from the LNs 24 h after SC MSC injection, and then added to CTV-labelled, anti-CD3/anti-CD28 activated splenic CD4+CD25− T cells at 1:5 ratio for 72 h. (B) Flow cytometric profile of CTV dilution (indicating T cell proliferation) in the presence of innate immune cells purified from LN of untreated mice (UNT LN) or LNs from SC MSC-injected mice (MSC LN). (C) Division index of T cell proliferation. Data expressed as mean ± SEM; n = 3 mice per group. (D) Representative flow cytometry plot showing Treg gating. (E) Relative change in Treg proportion in the presence of LN neutrophils, monocytes, DCs or macrophages after standardising to T cells alone group). Data expressed as mean ± SEM; n = 3 mice per group. (F) Draining (dLN; white bar) and non-draining LNs (ndLN; shaded bar) were harvested from mice that received SC injection of viable or apoptotic MSCs into the inflamed hock. Flow cytometry plots showing viability of MSCs treated with DMSO (mock) or BH3 mimetics prior to SC injection. (G) Treg cellularity in the dLN (white bars) and ndLNs (shaded bars) following LPS and SC MSC injection. (H) Changes in the PD-1 expression on Tregs after SC MSC injection, and Pearson correlation analysis of Treg PD-1 expression level and Treg proportion in the dLNs from LPS and MSC groups (Pearson r=-0.861, p = 0.00137). (I) CD44 expression on activated Tregs, and changes in CD44hi Tregs in dLNs and ndLNs after SC MSC injection. (J) Changes in PD-1 expression on CD44hiTregs after SC MSC injection, and Pearson correlation analysis of CD44hi Treg PD-1 expression level and Treg proportion in the dLNs from LPS and MSC groups (Pearson r=-0.883, p = 0.001). Data expressed as mean ± SEM; n = 5 mice per group. One-way ANOVA, Tukey’s multiple comparison, comparing UNT, LPS and MSC dLNs; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. (K) Treg cellularity in dLNs and ndLNs following SC injection with ApoMSC. (L) Changes in PD-1 expression on Tregs after SC injection with ApoMSC, and Pearson correlation analysis of Treg PD-1 expression level and Treg proportion in the dLNs from LPS and ApoMSC groups (Pearson r=-0.239, p = 0.392). Data expressed as mean ± SEM; n = 5–10 mice per group. One-way ANOVA, Tukey’s multiple comparison, comparing UNT, LPS and ApoMSC dLNs; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant
As MerTK+ macrophages from the LN of SC MSC-injected mice displayed increased IL-10 production and enhanced capacity for Treg expansion, we performed a more detailed analysis of the Treg population within the LN (Fig. 4F). There was a pronounced increase in Foxp3+ Tregs in the draining popliteal LN of MSC-treated mice (Fig. 4G). Strikingly, Tregs from SC MSC-injected mice displayed decreased PD-1 expression (Fig. 4H). PD-1 expression on Tregs has been shown to be inversely proportional to their suppressive activity [41, 42]. A recent study showed that PD-1 on Tregs inhibits Treg activation and suppressive activity, as PD-1-deficient Tregs are more immunosuppressive [43]. PD-1–deficient Tregs exhibit an activated phenotype (CD44hi CD62Llo) [43]. In line with this, there was an increase in activated CD44hi Tregs (Fig. 4I), which displayed decreased PD-1 expression levels (Fig. 4J), in the LN of SC MSC-injected mice. No significant difference in Tregs was observed in the non-draining LN of SC MSC-injected mice.
As the changes in Tregs were observed in the draining LN when the bulk of SC-injected MSCs were still viable at the site of injection (indicated by strong bioluminescent signals) 20 h after injection, we next investigated whether these changes were also induced by SC-injection of apoptotic MSCs (ApoMSCs). MSCs were treated with BH3-mimetic drugs to induce apoptosis via the intrinsic pathway [8] and mice received a SC injection of ApoMSCs in early apoptosis (Annexin V+PI−) directly in the inflamed hock. No changes in the number of Tregs (Fig. 4K), or their PD-1 expression (Fig. 4L), were observed in the LN following SC injection of ApoMSCs at this early timepoint. Thus, SC injection of viable MSCs but not ApoMSCs increased the number of activated CD44hi Tregs with decreased PD-1 expression in the draining LN.
SC MSC injection into inflamed skin inhibits the TNF response of LN neutrophils to LPS rechallengeHaving found that SC MSC injection promoted immunoregulatory cells in the LN draining the inflamed tissue, we next examined whether there was a suppression of the inflammatory response by analysing the production of pro-inflammatory TNF by LN cells following SC MSC injection. As MSCs were detectable up to 5 days after SC injection (Fig. 1B), a group of mice was injected with ApoMSCs as comparison, and the popliteal LN draining the inflamed tissue was analysed 4 h after LPS injection (Fig. 5A, B). Neutrophils were robust producers of TNF in response to LPS restimulation, with neutrophils accounting for ~ 40% of TNF-producing cells in the draining LN (Fig. 5C). Over 60% of neutrophils stained positive for intracellular TNF production (Fig. 5D). The popliteal LN from mice injected with viable MSCs showed a reduction in TNF-producing neutrophils upon restimulation with LPS (Fig. 5E, left panel). The reduction in TNF+ neutrophils was not observed in LN from mice that received ApoMSC (Fig. 5E, right panel). There were no significant changes in TNF+ monocytes with either viable MSC or ApoMSC injection (Fig. 5F, G). Further examination of neutrophils in the LN showed that whilst LPS injection increased their cell number and MHC class II expression, there were no significant changes with SC MSC injection (Fig. 5H). Thus, SC MSC injection decreased the TNF response of LN neutrophils to LPS, which required MSCs to be viable at the time of injection. The reduction in TNF was not due to a decrease in the number, or activation, of neutrophils in the LN.
Fig. 5
SC MSC injection inhibits TNF production by LN neutrophils in response to LPS re-exposure. (A) C57BL/6 mice were SC-injected with 30 ng LPS in 50 µl PBS in the left hock, and 1 h later 1 × 106 human adipose MSCs in the same hock. The popliteal LN draining the LPS-injected hock was harvested 4 h after LPS injection (i.e. 3 h after MSC treatment) and restimulated with 10 µg/ml LPS for 4 h in vitro. Intracellular TNF production was used as an indicator of the pro-inflammatory response to the LPS rechallenge. (B) Flow cytometry plots showing the gating strategies for identifying the TNF-producing (TNF+) populations after in vitro restimulation of bulk LN cells with LPS. (C) Representative flow cytometry plots showing the gating for TNF+ neutrophils. (D) Number of TNF+ neutrophils after LPS restimulation of LN cells from mice that received SC injection of viable MSCs (left panel) or ApoMSC (right panel). Data expressed as mean ± SEM; n = 5–10 mice per group from 2 independent experiments. One-way ANOVA, Tukey’s multiple comparison; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. E) Representative flow cytometry plots showing the gating for TNF+ monocytes. (F) Number of TNF+ monocytes after LPS restimulation of LN from mice that received SC injection of viable MSCs (left panel) or ApoMSC (right panel). Data expressed as mean ± SEM; n = 5–10 mice per group from 2 independent experiments. One-way ANOVA, Tukey’s multiple comparison; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. (G) Cellularity and expression of MHCII, CD86 and PD-L1 on neutrophils in the dLN after SC MSC injection. Data expressed as mean ± SEM; n = 5 mice per group. One-way ANOVA, Tukey’s multiple comparison; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant
Direct MSC injection into the inflamed skin is necessary for immunoregulatory effects in the LNThe immunomodulatory effects induced by SC MSC injection were observed in the LN draining the inflamed hock, but not in the non-draining LN. We therefore investigated whether SC MSC injection into the non-inflamed hock would induce the same effects. Using the above LPS-induced inflammation model, we first confirmed that LPS injection into the hock led to an increase in pro-inflammatory cytokines TNF, IL-1β, IL-6 and MCP-1 in the skin from the inflamed hock and did not induce any cytokine changes on the contralateral side (non-inflamed hock) (Fig. 6A). We then repeated the bioluminescence imaging to determine whether local inflammation affects the in vivo persistence of SC-injected MSCs. Fluc-GFP+ MSCs were injected via the SC route into the inflamed skin area (Fig. 6B) or the non-inflamed contralateral hock (Fig. 6C) 1 h post-LPS injection. Mice were imaged 10 min after SC MSC injection, 3 h later and then daily. Local inflammation did not increase the bioluminescent signals of fluc-GFP+ MSCs, neither did inflammation at a distance from MSC injection drive cell migration beyond the injection site (Fig. 6B, C). The bioluminescent signals lasted around 5 days, with a slight increase in bioluminescence signal during the first 2 days (Fig. 6D). Direct imaging of the popliteal LNs did not show drainage of significant numbers of MSCs to the local popliteal LN (data not shown). Although unlikely, we cannot rule out the potential dissemination of small numbers of MSCs, which were below the detection range, to other sites of the body.
Fig. 6
Contralateral SC MSC injection does not inhibit LN neutrophil TNF response to LPS re-exposure. (A) C57BL/6 mice were SC-injected with 30 ng LPS in 50 µl PBS in the left hock, and 4 h later the skin around the LPS injection site (LPS inflamed) and on the contralateral hock (LPS non-inflamed) was excised and homogenised for cytokine analysis. Data expressed as mean± SEM; n = 10 mice per group from 2 independent experiments. One-way ANOVA, Tukey’s multiple comparison, * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. (B) Bioluminescent imaging of mice that received SC injection of 30 ng LPS in 50 µl PBS in the left hock, and 1 h later 1 × 106 luciferase-expressing MSCs in the same hock, or (C) in the non-inflamed hock on the contralateral side. (D) Bar graphs represent changes in radiance over time. n = 3 mice per group from one experiment. (E) C57BL/6 mice were SC-injected with 30 ng LPS in 50 µl PBS in the left hock. 1 h after LPS injection, mice received a SC injection of 1 × 106 human adipose MSCs (MSC-c) or control PBS treatment (PBS-c) in the contralateral (non-inflamed) hock. The popliteal LNs draining the LPS-injected and the contralateral skin were harvested 4 h after LPS injection (i.e. 3 h after MSC-c/PBS-c treatment) and restimulated with 10 µg/ml LPS for 4 h. Intracellular TNF production was used as an indicator of the pro-inflammatory response to the LPS rechallenge. (F) Number of TNF+ neutrophils after LPS restimulation of LN cells from mice that received SC injection of MSC or control PBS on the contralateral hock. Data expressed as mean ± SEM; n = 5 mice per group. One-way ANOVA, Dunnett’s multiple comparison; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant. (G) Number of TNF+ monocytes after LPS restimulation of LN cells from mice that received SC injection of MSC or control PBS on the contralateral hock. Data expressed as mean ± SEM; n = 5 mice per group. One-way ANOVA, Dunnett’s multiple comparison; * p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant.
We next investigated whether contralateral SC MSC injection into the non-inflamed hock could remotely regulate the inflammatory response to LPS within the draining LN (Fig. 6E). Analysis of the popliteal LN draining the inflamed hock 4 h after LPS injection showed that neither contralateral MSC injection nor PBS control treatment had an effect on the number of TNF-producing neutrophils or monocytes in response to LPS restimulation (Fig. 6F, G; inflamed). There was also no change in the number of TNF+ neutrophils or monocytes in the popliteal LN from the non-inflamed hock in any of the treatment groups (Fig. 6F, G; non-inflamed). Thus, in contrast to direct MSC injection into inflamed tissue, contralateral MSC injection did not affect the LN inflammatory response to LPS. This finding indicates that ‘priming’ of SC-injected MSCs by inflammatory cytokines in the local milieu is important for their immunoregulatory effects in the LN.
Taken together, our data demonstrate that SC injection of MSCs into inflamed tissue induced an increase in immunoregulatory cells in the tissue-draining LN and reduced the responsiveness of neutrophils to subsequent inflammatory challenge. In contrast to IV delivery of MSCs, the immunoregulatory effects of SC MSC injection on the local draining LN are not coupled with MSC apoptosis.
留言 (0)