
記住我
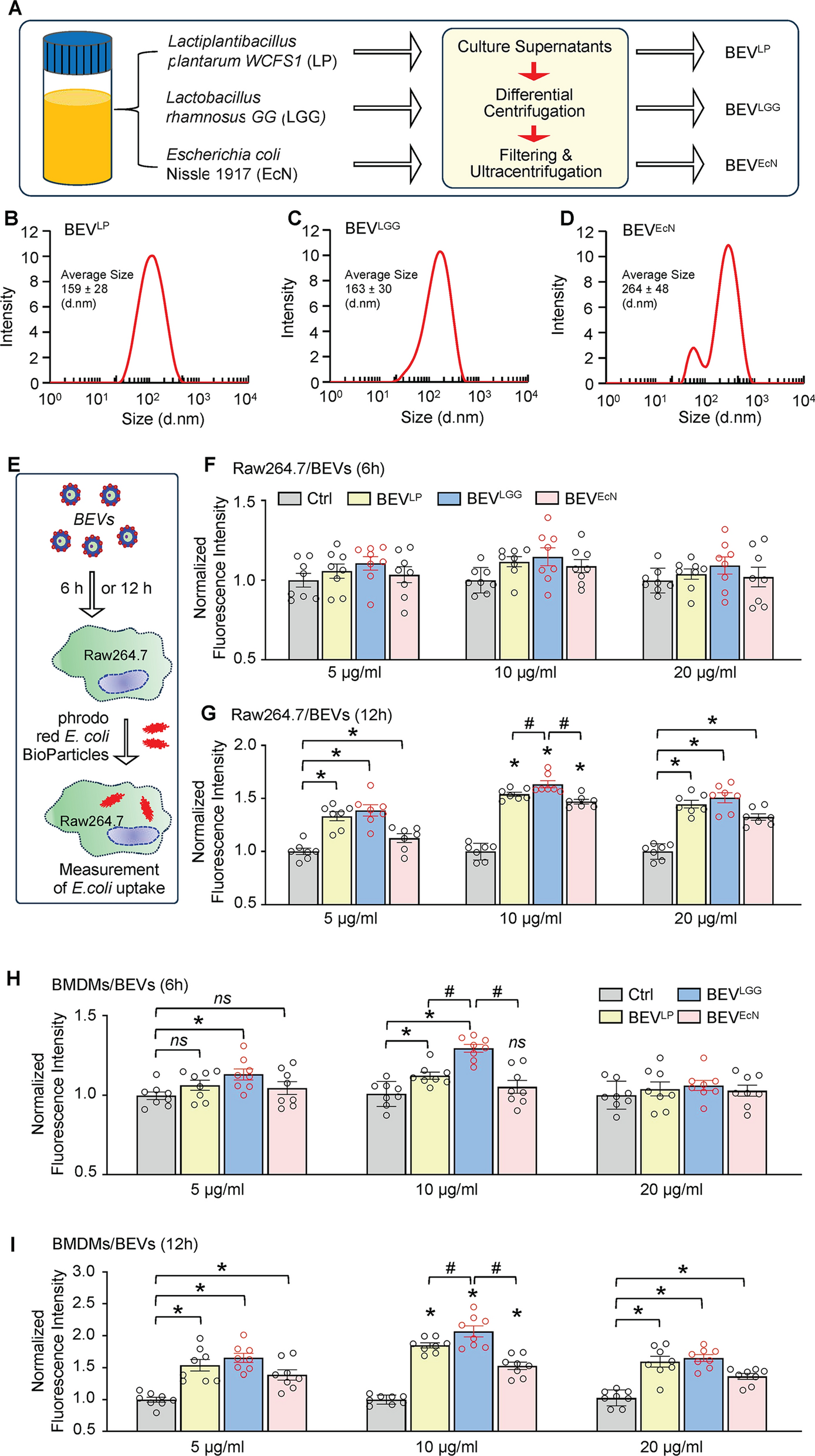



Given that LP WCFS1, LGG and EcN are three well-characterized probiotics, we cultured them under microaerobic conditions and collected the culture supernatants to isolate the bacteria-released extracellular vesicles (BEVs), as depicted in Fig. 1A. Using a protein concentration assay and NTA, we observed that the protein content of the BEVLP preparations was approximately 0.91 µg/µl, and the average diameter of the BEVLP was 159 ± 28 nm (Fig. 1B). Moreover, BEVLGG contained 1.02 µg/µl protein with an average diameter of 163 ± 30 nm (Fig. 1C), and BEVEcN contained approximately 1.17 µg/µl protein with an average diameter of 264 ± 48 nm (Fig. 1D).
Fig. 1
Physical characterization of BEVs released from probiotic bacteria and their effects on macrophage phagocytosis. A Flow chart for the collection and purification of BEVs from the cultivated probiotic bacteria. B-D Size distribution [B: LP WCFS1, C: LGG, D: EcN] of these BEVs determined by nanoparticle tracking analysis. E A cartoon scheme describing the procedure to measure phagocytic capacity in Raw264.7 macrophages after treatment with BEVs. F-I After incubation of Raw264.7 macrophages [F: 6 h, G: 12 h] and BMDMs [H: 6 h, I: 12 h] with BEVs derived from three probiotics at different doses for 6 h and 12 h, respectively, the phagocytic capacity was measured by determining the red fluorescence intensity with a plate reader. All results are presented as mean ± SEM and analyzed by 2-way ANOVA (*p < 0.05 vs. control group, #p < 0.05 vs. BEVLP and BEVEcN, ns: non-significance)
Considering that bacteria can naturally stimulate macrophage phagocytosis, we next asked whether these bacteria-released EVs had a similar effect on macrophages. To this end, we initially performed bacterial phagocytosis assays using a cell line, Raw264.7 macrophages. As depicted in Fig. 1E, BEVLP, BEVLGG, or BEVEcN were added to Raw264.7 cells at different doses (5, 10, and 20 µg/mL), and the cells were incubated for 6 h or 12 h. The phagocytic capacity was subsequently assessed by measuring the uptake of red fluorescent-conjugated pHrodo E. coli BioParticles. Interestingly, after incubating for 6 h, neither of these BEVs promoted the uptake of pHrodo E. coli BioParticles by Raw264.7 cells, as evidenced by the similar red fluorescence intensities among these BEV-treated cells and control cells (Fig. 1F). However, after the incubation time was extended to 12 h, all three BEV-treated Raw264.7 cells displayed a significantly higher intercellular red fluorescence intensity than did the control-treated cells (Fig. 1G). Further analysis of Raw264.7 cells revealed that incubating BEVLGG at a dose of 10 µg/mL for 12 h resulted in greater phagocytic capacity than incubating either BEVLGG- or BEVEcN-treated cells under the same conditions (Fig. 1G). These results indicate that BEVs are capable of stimulating macrophage phagocytosis but have strain-, time-, and dose-dependent effects.
To further determine whether BEVs could play a similar role in ex vivo macrophages as in vitro cell line above, we isolated/cultured BMDMs and incubated them with these BEVs at the same doses and duration as those used for Raw264.7 cells. Unlike their effects on Raw264.7 cells, different bacterial strain-derived EVs have distinctive phagocytosis capacities on BMDMs. As shown in Fig. 1H, after incubation with 5 µg/mL BEVs for 6 h, only BEVLGG-treated BMDMs exhibited a significantly higher uptake of E. coli bioparticles, compared to control-treated cells. Importantly, BMDMs were treated with 10 µg/mL BEVs for 6 h, both BEVLP and BEVLGG, but not BEVEcN, were able to augment macrophage phagocytosis compared to that in control-treated cells. (Fig. 1H). Notably, BEVLGG could stimulate macrophage phagocytosis to a greater degree than BEVLP. Unexpectedly, when the BEV dose was increased to 20 µg/mL for 6 h, none of the three BEVs promoted macrophage phagocytosis (Fig. 1H). Consistently, when extended BEV-treatment time to 12 h, all BEVs could significantly enhance phagocytosis capacity in BMDMs, compared to control cells (Fig. 1I), similar to what was observed in the Raw264.7 cells (Fig. 1G). Notably, among the three bacterial strain-derived EVs, BEVLGG had the greatest capacity to stimulate BMDMs phagocytosis at a dose of 10 µg/mL (Fig. 1I). Therefore, we selected probiotic LGG to generate BEVs and chose a dose of 10 µg/mL for the following in vitro studies.
Characterization of BEVs released from LGG cultured under acid, neutral, and alkaline conditions and their effects on macrophage phagocytosisIt has been reported that pH value, an important parameter of culture medium, directly affects bacterial growth, metabolism, and synthesis of bioactive substances (Wang et al. 2020; Zaghloul et al. 2023). Accordingly, the properties of BEVs collected from probiotics under different pH conditions may be distinctive. Along this line, we cultured LGG under acid (pH5), standard (pH6.5), and alkaline (pH8) conditions to generate BEVpH5, BEVpH6.5, and BEVpH8, as depicted in Fig. 2A. Firstly, we measured LGG growth rate and observed that three pH conditions did not significantly alter their growth curve (Supplementary Fig. S1A). Given that LTA is a major component of the cell wall of Gram-positive bacteria and plays a role in bacterial adherence, immune system activation, and inflammation, we then measured LTA levels in these BEVs. Unexpectedly, our results showed that the LTA contents in BEVpH5, BEVpH6.5, and BEVpH8 were very low and less than 0.45 pg/µg, and there were no statistical differences among the three groups of BEVpH5, BEVpH6.5, and BEVpH8 (Supplementary Fig. S1B). However, the average diameter size of BEVpH5 was significantly larger (211 ± 39 nm; Fig. 2B), compared to that of BEVpH6.5 (159 ± 28 nm; Fig. 2C) and BEVpH8 (157 ± 30 nm; Fig. 2D).
Fig. 2
Physical characterization of BEVs released from LGG cultured under acidic, neutral, and alkaline conditions and their effects on macrophage phagocytosis. A Flow chart for the isolation of BEVs from LGG cultivated under three different pH conditions. B-D Size distribution [B: BEVpH5, C: BEVpH6.5, D: BEVpH8] of these BEVs determined by nanoparticle tracking analysis. E-J Representative confocal images of E. coli bioparticles engulfed by Raw264.7 macrophages (E) and BMDMs (H) after treatment with LGG-derived BEVs (10 µg/mL) (scale bar, 10 μm). The normalized mean fluorescence intensity in Raw264.7 macrophages (F) and BMDMs (I) was quantified. After incubation of Raw264.7 macrophages (G) and BMDMs (J) with BEVs derived from LGG cultured under three different pH conditions for 12 h, the phagocytic capacity was assessed by determining the red fluorescence intensity with a plate reader. K-N Representative flow cytometry histograms and median fluorescence intensity values showing phagocytosis of E. coli bioparticles by Raw264.7 macrophages (K, L) and BMDMs (M, N) treated with LGG-derived BEVs cultured under three different pH conditions. All results are presented as mean ± SEM and analyzed by one-way ANOVA (*p < 0.05 vs. control group, #p < 0.05 vs. BEVpH5 and BEVpH6.5)
Next, we determined the effects of these BEVs on macrophage phagocytosis, using three different methods including: (1) confocal microscopy, (2) microplate reader, and (3) flow cytometry to measure fluorescence intensity of red pHrodo E. coli BioParticles taken up by macrophages. To exclude possible effects of LTA contained in the BEVs, we used PBS plus an equal amount of LTA that was included in the corresponding BEVLGG as a control. As shown in Fig. 2E/F, treatment of Raw264.7 macrophages with BEVpH5, BEVpH6.5, or BEVpH8 for 12 h significantly increased the uptake of red E. coli bioparticles, compared with control cells. Importantly, the highest amount of red bioparticles were observed in the BEVpH8-treated Raw264.7 cells, compared to both BEVpH5- and BEVpH6.5-treated groups as determined via confocal microscopy. Consistently, using microplate reader, we found that the intracellular red fluorescence intensity was significantly higher in all BEV-treated Raw264.7 macrophages than control-treated cells with the highest in BEVpH8-treated group (Fig. 2G). Similar results were also observed in BMDMs upon exposure to these BEVs for 12 h, followed by addition of red pHrodo E. coli BioParticles and subsequently analyzed by confocal microscopy (Fig. 2H/I) and microplate reader (Fig. 2J). Additionally, flow cytometry analysis revealed higher red fluorescence content in all BEV-treated Raw264.7 macrophages (Fig. 2K/L) and BMDMs (Fig. 2M/N), compared to the control-treated cells, while the red fluorescence intensity was strongest in BEVpH8-treated macrophages (Fig. 2K–N). Taken together, these data consistently demonstrate that BEVs released by LGG bacteria, regardless of culture pH conditions, can greatly promote macrophage phagocytosis. Despite that, BEVs from LGG cultured in pH8-MRS medium are the best at stimulating macrophage phagocytosis.
LGG-released BEVs enhance phagocytic and bactericidal activities of macrophagesTo further determine the effects of LGG-released BEVs on the control of bacterial dissemination, we next utilized live E. coli in place of fluorescence-conjugated bioparticles to infect BEV-treated macrophages, followed by bacterial uptake and intracellular killing assays. As shown in Fig. 3A/B, a greater number of bacteria was observed in all BEV-treated Raw264.7 cell lysates collected 30 min after bacterial infection, compared to the control group, while pre-treatment of Raw264.7 macrophages with BEVpH8 displayed the highest among three BEV-treated groups. These data further validate that LGG-BEVs can stimulate macrophages to take up bacteria with BEVpH8 to the greatest extent. At 4 h post-bacterial infection, we collected macrophage lysates and determined the number of bacterial retained in BEV-treated macrophages to calculate the percentage of bacteria killed according to the formula described in the Methods section. While there was no difference in the number of bacterial CFUs among the 4-h-cell lysates (Fig. 3A/C), the percentage of intracellular bacterial killing revealed that pre-treatment of Raw264.7 cells with BEVpH8 exhibited the highest bactericidal effects (83.1%), followed by pre-treatment with BEVpH6.5 (78.9%) and BEVpH5 (74.5%), all were better than control group (55.2%) (Fig. 3D). Similarly, BEV-treated BMDMs showed a significant increase in bacterial uptake and killing rate compared with control-BMDMs, meanwhile the most increase was observed in BEVpH8-treated BMDMs, compared to both BEVpH5- and BEVpH6.5-treated cells (Fig. 3E–H). Collectively, these data further indicate that probiotic LGG-released BEVs can remarkably enhance macrophage phagocytosis and bactericidal activity. Particularly, BEVs collected from pH8-condition LGG demonstrated the highest potency in promoting bacterial clearance in macrophages. Therefore, we selected BEVpH8 to test their therapeutic effects in septic mice (below).
Fig. 3
LGG-derived BEVs cultured under three different pH conditions can promote the phagocytic activity and intracellular bactericidal ability of macrophages toward live bacteria. Gentamicin protection assay was used to test phagocytic and bactericidal activities of Raw264.7 macrophages (A) and BMDMs (E) using live E. coli. Gentamicin (100 µg/mL) was added to the culture medium 1 h after infection with live E. coli (MOI = 20). After 30 min, the cell lysate was diluted using the serial dilution method and plated on LB agar plates. The CFUs were measured after 24 h as an indicator of phagocytosis capacity in Raw264.7 macrophages (B) and BMDMs (F). In addition, CFUs in Raw264.7 macrophages (C) and BMDMs (G) lysates were determined 4 h after the addition of gentamicin to assess the number of bacteria remained within the macrophages. The killing percentages of Raw264.7 macrophages (D) and BMDMs (H) were calculated as described in the Materials & Methods section (n = 6). All results are presented as mean ± SEM and analyzed by one-way ANOVA (*p < 0.05 vs. control group, #p < 0.05 vs. BEVpH5 and BEVpH6.5)
Administration of BEVpH8 attenuates polymicrobial sepsis-induced mortality and organ injury in miceTo investigate the in vivo therapeutic effects of BEVpH8, we employed a feces-injection-in-peritoneum (also named fecal-induced peritonitis, FIP) model of murine sepsis, which is closely mimicked the clinical conditions (Sharma et al. 2023) and more suitable for standardization than cecal ligation and puncture (CLP)-sepsis model (Fang et al. 2020). At 1-h post-FIP, BEVpH8 (2 µg/g body weight) or control (PBS plus the same amount of LTA included in the BEVs, 0.6 pg/g body weight) was administered via R.O. injection followed by monitoring for mortality over a 3-day period and other experimental analysis (Fig. 4A). We observed that BEVpH8-treated mice exhibited a significantly higher survival rate than control-treated mice did over a period of 3 days post-FIP (Fig. 4B). Additionally, we recorded the MSS, a quantitative metric in which behavior, appearance, activity level, respiratory rate, and other physiological indicators are scored. We found that sepsis scores at various time points were lower in the BEVpH8-treated group compared to the control group (Fig. 4C). This suggests that the control group exhibited more severe sepsis-related symptoms and organ dysfunction after FIP. Notably, there was a decreasing trend in the murine sepsis scores in the BEVpH8-group starting at 8 h post-FIP, indicating effective control of the bacterial infection. Furthermore, we observed that FIP-induced lung injury was significantly mitigated in BEVpH8-treated group, as evidenced by the greater decrease in neutrophil infiltration, formation of hyaline membranes, thickness of alveolar wall, and the alveolar collapse in lung tissues collected from BEVpH8-injected mice (injury score: 0.358 ± 0.064), compared to those from control-treated mice (injury score: 0.608 ± 0.091) (Fig. 4D/E). Next, we measured the serum levels of alanine aminotransferase (ALT, biomarker of liver injury), aspartate aminotransferase (AST, biomarker of liver and heart injury), creatinine (Cr, biomarker of kidney injury), and lactate dehydrogenase (LDH, biomarker of tissue damage) in BEV-treated mice at 12 h post-FIP, in comparison with control-treated septic mice. As shown in Fig. 4F–I, the serum concentrations of these biomarkers were significantly decreased in BEVpH8-treated mice, compared to control mice. Collectively, these data suggest that the administration of BEVpH8 could significantly rescue FIP-induced multi-organ injury in mice.
Fig. 4
Administration of BEVpH8attenuates polymicrobial sepsis-induced mortality and organ injury in mice. A Graphic illustration of experimental design: Feces-injection-in-peritoneum (FIP)-induced sepsis mouse model, and administration of BEVpH8 (2 µg/g), or control through the retro-orbital (R.O.) injection at 1 h post-FIP. B Kaplan–Meier survival curves were generated to compare mortality between groups, and significance was determined by log-rank (Mantel-Cox) test (*p < 0.05). C The murine sepsis score (MSS) was assessed at various time points by observing and recording behavioral, visual, activity levels, respiratory rate, and other physiological parameters of the mice (*p < 0.05). D Representative images of lung sections stained with hematoxylin and eosin (H&E) from sham, control-, and BEVpH8-treated mice at 12 h post-FIP (scale bar, 50 μm). E The lung injury scores were assessed as described in the Materials & Methods section (n = 6). F–I Serum levels of ALT (F), AST (G), Cr (H), and LDH (I) in each group were measured at 12 h post-FIP using ELISA kits (n = 6). All results are presented as mean ± SEM and analyzed by 2-way ANOVA (*p < 0.05 when compared with respective control group)
Administration of BEVpH8 suppresses inflammatory response and reduces bacterial burden in FIP-septic miceGiven that bacterial burden and inflammatory responses are important driving factors for the sepsis-triggered organ injury, we then collected peripheral blood and PLF from septic mice at 6 h and 12 h after FIP to determine the impact of BEVpH8 on the systemic and local inflammatory response and bacterial clearance. Using ELISA kits, we measured the concentrations of inflammatory cytokines and chemokines such as IL-1β, IL-6, TNF-α, and MCP-1, in the blood and PLF collected above. Our results show that the serum levels of IL-1β (Fig. 5A), IL-6 (Fig. 5B), TNF-α (Fig. 5C), and MCP-1 (Fig. 5D) were significantly lower in BEVpH8-treated mice than in control-treated mice at 6 h post-FIP and were more pronounced at 12 h post-FIP. Similarly, BEVpH8-mediated suppression of inflammatory response was also observed in the PLF (Fig. 5E–H). Additionally, we assessed mRNA levels of inflammatory cytokines and chemokines in lung tissues at 12 h post-FIP. Our results demonstrated that mRNA levels of IL-1β (Supplementary Fig. S2A), IL-6 (Supplementary Fig. S2B), TNF-α (Supplementary Fig. S2C), and MCP-1 (Supplementary Fig. S2D) in the lungs of BEVpH8-treated group were significantly lower, compared to the control group.
Fig. 5
Administration of BEVpH8 mitigates both local and systemic inflammatory responses A, E IL-1β, B, F IL-6, C, G TNF-α, and D, H MCP-1 levels in the serum and PLF of Sham, control- and BEVpH8-treated mice at 6 h and 12 h post-FIP. All results are presented as mean ± SEM and were compared to respective control group at each time point by 2-way ANOVA (n = 6, *p < 0.05 when compared with respective control group)
Next, we evaluated bacterial clearance in various organs, including the lung, spleen, liver, kidney, and heart, by measuring the expression levels of 16S rRNA, a marker of bacterial presence. We detected that the levels of bacterial 16S rRNA were significantly decreased in the lungs (Fig. 6A), spleen (Fig. 6B), liver (Fig. 6C), and kidneys (Fig. 6D), collected from BEVpH8-treated mice, compared to those organs from control-treated mice at 6 h post-FIP, which were more dramatically reduced at 12 h post-FIP (Fig. 6A–D). However, there was much less bacterial 16S rRNA detected in the heart which showed no significant difference between two groups at either 6 h or12 h post-FIP (Supplementary Fig. S3A). Lastly, the bacterial load was examined in the blood and PLF of septic mice. We observed that bacterial burden was remarkably reduced in both blood (Fig. 6E, F) and PLF (Fig. 6G, H) collected from BEVpH8-treated mice in comparison with control-treated mice at 6 h and 12 h post-FIP. It is important to note here that the bacterial load was highly increased in the PLF compared to the blood, particularly at 12 h post-FIP. This phenomenon could be attributed to the feces-injection i.p. where bacteria continue to proliferate to peak levels in the peritoneal cavity at 6–12 h after FIP. Taken together, these data indicate that BEVpH8 has the strong potential to limit both systemic and local inflammatory responses as well as bacterial dissemination during sepsis.
Fig. 6
Administration of BEVpH8 decreases bacterial burden in multiple organs, blood, and PLF after FIP challenge. A–D Relative expression level of bacterial 16S rRNA in organs [A: Lung, B: Spleen, C: Liver, D: Kidney] of control- and BEVpH8-treated mice at 6 h and 12 h post-FIP by RT-qPCR (n = 6). E–H The bacterial burden in both blood (E, F) and PLF (G, H) were measured in control- and BEVpH8-treated mice at 6 h, and 12 h post-FIP. All results are presented as mean ± SEM and analyzed by 2-way ANOVA (*p < 0.05 when compared with respective control group)
RNA sequencing analysis of gene expression profiles of BEV-treated BMDMsTo elucidate the potential mechanisms by which LGG-derived BEVs enhance macrophage phagocytosis of bacteria, we treated BMDMs with BEVpH5, BEVpH6.5, BEVpH8, or control for 12 h, followed by isolating total RNAs for RNA-seq. Given that BEVpH8 has the strongest capacity in stimulating macrophage phagocytosis, then we plotted heatmaps and volcano plots for three comparisons as BEVpH8 vs. Control (Fig. 7A/B), BEVpH8 vs. BEVpH5 (Fig. 7C/D), and BEVpH8 vs. BEVpH6.5 (Fig. 7E/F). Interestingly, among the top 30 differentially expressed genes, we were surprised to find that only three macrophage receptor genes including FPR1, FPR2, and MARCO were dramatically and commonly upregulated in BEVpH8-treated BMDMs, compared to control-, BEVpH5-, and BEVpH6.5-cells, respectively (Fig. 7A–F). FPR1 and FPR2 are formyl peptide receptors (FPRs) that are expressed at low levels on the surface of macrophages under basal conditions, while they can recognize bacterial peptides and initiate phagocytosis (He and Ye 2017; Liang et al. 2020). The macrophage receptor with collagenous structure (MARCO), a scavenger receptor, facilitates the recognition and uptake of microbial pathogens by macrophages (Zhou et al. 2024). Therefore, we selected these three receptor genes for validation via RT-qPCR. As shown in Fig. 7G, treatment of BMDMs with BEVpH5, BEVpH6.5, and BEVpH8 markedly upregulated the expression of FPR1, FPR2, and MARCO by > 2200-fold, > 96-fold, and > 128-fold, respectively, compared to control-treated control cells. Notably, BEV-mediated elevation of FPR1, FPR2, and MARCO expression was more pronounced in BEVpH8-BMDMs than in both BEVpH5- and BEVpH6.5-treated groups (Fig. 7G). Similar findings were also observed in Raw264.7 macrophages treated with BEVpH5, BEVpH6.5, and BEVpH8 (Fig. 7H).
Fig. 7
Gene expression profile in BMDMs treated with BEVs collected from the LGG cultures by high-throughput RNA sequencing. A–F Heatmap and volcano plot of the differentially expressed genes in BMDMs treated with control, BEVpH5, BEVpH6.5 or BEVpH8 [A, B: BEVpH8 vs. control; C, D: BEVpH8 vs. BEVpH5; E, F: BEVpH8 vs. BEVpH6.5] (n = 3). The expression of FPR1, FPR2, and MARCO were among the most significantly altered genes in BEV-treated BMDMs, which were further validated in BMDMs (G) and Raw264.7 macrophages (H) by RT-qPCR (n = 6). I, J ROS levels were measured in BMDMs (I) and Raw264.7 macrophages (J) upon treatment with BEVLGG for 12 h, followed by incubation with heat-killed E. coli for 30 min. K, L BMDMs were pretreated with control, BEVpH5, BEVpH6.5 or BEVpH8 (10 µg/mL) for 12 h. Then, cells were collected for Western blot analysis of total and phosphorylated Akt. Representative immunoblots (K) and their quantification analysis results (L) showing Akt activation. GAPDH was used as a loading control (n = 5). All results are presented as mean ± SEM and analyzed by one-way ANOVA (*p < 0.05 vs. controls, #p < 0.05 vs. BEVpH5 and BEVpH6.5)
Currently, it is well recognized that upregulation of FPR1/FPR2 can activate downstream signaling cascades, including: (1) Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, which induces ROS generation to facilitate bacterial killing inside macrophages (Xu et al. 2016); and (2) PI3K-Akt which can promote MARCO expression (Sun et al. 2023). Therefore, we next measured ROS levels in these BEV-treated macrophages upon stimulation with heat-killed E. coli. Our results showed that BEVpH8-treated BMDMs contained the highest levels of ROS, compared with other two BEV-treated groups, while all BEV-treated cells had higher levels of ROS than control-treated cells (Fig. 7I). Similar phenomena were also found in Raw264.7 macrophages treated with these LGG-derived BEVs (Fig. 7J). Regarding the PI3K-Akt pathway, we assessed Akt activity by measuring its phosphorylation in BEV-treated or control-treated BMDMs, using Western-blotting analysis. As shown in Fig. 7K/L, the phosphorylated Akt levels were significantly increased in all BEV-treated groups with the highest in BEVpH8-treated BMDMs, compared to control-treated BMDMs. Collectively, these data consistently indicate that BEVs released by probiotic LGG, especially cultured under pH8 condition can stimulate macrophages to strongly upregulate FPR1/2 expression, leading to increased ROS production and Akt activity as well as MARCO expression.
Inhibition of FPR1/2 attenuates BEVpH8-mediated effects on macrophage phagocytosisLastly, we asked whether BEVpH8-mediated augmentation of macrophage phagocytosis is dependent on the upregulation of FPR1/2. To address this question, we pre-treated macrophages (Raw264.7 cells and BMDMs) with Boc2, a specific antagonist of FPR1/2, for 2 h, followed by incubation with BEVpH8 for 12 h and subsequently, macrophage phagocytosis analysis was performed using red pHrodo E. coli BioParticles. As shown in Fig. 8A, B, BEVpH8-treated Raw264.7 cells took up a remarkably higher amount of red E. coli particles than control-treated cells, which is consistent with the above data in Fig. 2E, F. However, BEVpH8-mediated dramatic increase of phagocytosis was greatly attenuated by pre-treatment of Raw264.7 cells with FPR1/2 inhibitor, Boc2; albeit it remained significantly higher than control cells (Fig. 8A, B). These results were further validated by measuring red fluorescence intensity using 96-well plates (Fig. 8C). Together, our data suggest that BEVpH8-mediated augmentation of phagocytosis is dependent on FPR1/2, at least in part. Similar outcomes were also shown in BMDMs upon pre-treated with Boc2, followed by addition of BEVpH8 for 12 h (Fig. 8D–F). Accordingly, further analysis using live bacterial infection revealed that the CFU counts from cell lysates of BEVpH8-treated Raw264.7 cells at 30-min post-infection were increased by 1.7-fold than controls in the absence of Boc2, but only increased by 0.3-fold than controls in the presence of Boc2 (Fig. 8G and Supplementary Fig. S4A). This suggest that BEVpH8-mediated uptake of bacteria is greatly blocked in macrophages when FPR1/2 are inhibited by Boc2. Although there were no difference in the CFU counts from all groups of cell lysates collected at 4-h post-infection (Fig. 8H), the percentage of intracellular bacterial killing was remarkably higher in BEVpH8-treated Raw264.7 cells, compared to control cells without pre-inhibition of FPR1/2 by Boc2 (Fig. 8I), but such BEVpH8-induced increase of bacterial killing rate was significantly diminished in Raw264.7 cells upon pre-treatment with Boc2 (Fig. 8I). In addition, similar inhibitory effects of Boc2 on BEVpH8-induced phagocytosis and bacterial clearance were also observed in BMDMs (Fig. 8J-L and Supplementary Fig. S4B). Collectively, our results demonstrate that inhibition of FPR1/2 could largely attenuate BEVpH8-mediated boost effects on macrophage phagocytosis and bactericidal activity.
Fig. 8
Inhibition of FPR1/2 abolishes BEVpH8-mediated effects on macrophage phagocytosis and bacterial killing activities. Raw264.7 macrophages (A-C) and BMDMs (D-F) were pre-treated with Boc2 (N-tert-butyloxycarbonyl-Phe-Leu-Phe-Leu-Phe), a specific antagonist of FPR1/FPR2 for 2 h, followed by incubation with BEVpH8 for 12 h. Then phagocytic capacity was assessed by the uptake of red E. coli bioparticles. Representative confocal images of phagocytosis of E. coli bioparticles by Raw264.7 macrophages (A) and BMDMs (D) (scale bar, 10 μm). The normalized mean fluorescence intensity in Raw264.7 macrophages (B) and BMDMs (E) was quantified. The phagocytic capacity of Raw264.7 macrophages (C) and BMDMs (F) was assessed by measuring the red fluorescence intensity using a plate reader. Gentamicin protection assay was used to detect the phagocytic ability and bactericidal activities in these Raw264.7 macrophages (G-I) and BMDMs (J-L) after treatment as described above. Gentamicin (100 µg/mL) was added to the culture medium 1 h after infection with live E. coli (MOI = 20). After 30 min, cell lysate was extracted with serial dilution, then plated on LB agar plates. The CFUs were measured as an indicator for phagocytosis capacity of Raw264.7 macrophages (G) and BMDMs (J). CFUs were determined in lysates of Raw264.7 macrophages (H) and BMDMs (K) at 4 h after the addition of gentamicin to assess the number of bacteria remained within macrophages. The killing percentages of Raw264.7 macrophages (I) and BMDMs (L) were calculated as described in the Materials & Methods section (n = 6). All results are presented as mean ± SEM and analyzed by 2-way ANOVA (*p < 0.001, #p < 0.05)
留言 (0)