
記住我
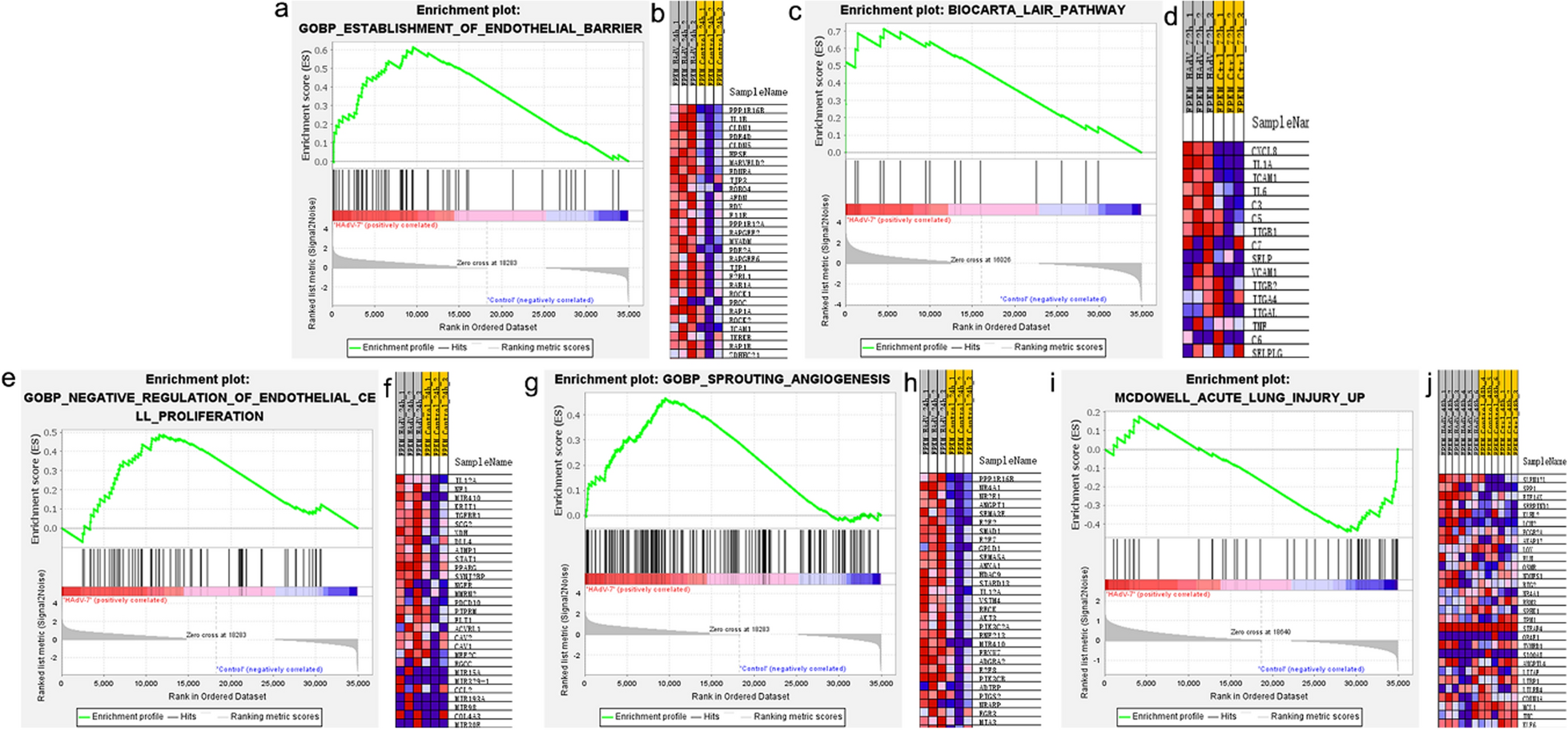
The initial experiments were carried out to determine the tolerance of cell cultures to ECSN6. The cell cultures were treated apically twice a day for up to 96 h with placebo or varying concentrations of ECSN6 and examined for TER, IL-8 and LDH every 24 h. There was no significant difference between PBS- and placebo-treated cultures in TER, IL-8 expression and LDH release indicating that placebo does not have any toxic or pro-inflammatory effects on epithelial cells. Compared to placebo or PBS, treatment with 20% ECSN6 gradually enhanced TER up to 72 h and was significantly increased at 72 h. In 40% ECSN6 treated cells, the TER significantly increased starting from 24 h without increasing IL-8 expression or LDH release (Supplemental Fig. 1A-1C). These results show that ECSN6 improves barrier integrity and function in non-infected cultures. Beyond 40%, ECSN6 caused cell death and IL-8 release. Cell cultures treated with 10% and 20% ECSN6 for 72 h showed increased localization of ZO-1 and E-cadherin at the intercellular junctions (Supplemental Fig. 2). Compared to cells treated with 10% or 20% ECSN6, cells treated with ECSN6 to 40% for 72 h showed slight reduction in localization of both ZO-1 and E-cadherin to the intercellular junctions and had an elongated shape. These results suggest that 20% ECSN6 may improve TER by increasing the expression of tight and adherence proteins or enhancing their localization to the intercellular junctions. Therefore, in the subsequent experiments we used 20% ECSN6.
Treatment with ECSN6 prevents RV-A1-induced reduction in TER and attenuates permeability to inulinMucociliary-differentiated cultures infected apically with sham or RV-A1 at MOI of 1 were treated with placebo or 20% ECSN6 twice a day starting from 2 h post-infection. Treatment with 20% ECSN6 increased TER in sham and RV-A1-infected cultures at all time points (Fig. 1A and C). RV-A1-infected/placebo-treated cultures showed reduction in TER at 24 h post-infection which returned to baseline at 48 h post-infection. Treatment with ECSN6 completely prevented the effect of RV-A1 on TER.
Fig. 1
ECSN6 prevents RV-A1-induced reduction in TER and reduces permeability to inulin. Mucociliary-differentiated airway epithelial cultures were infected apically with RV-A1 or sham. The cultures were treated with placebo or 20% ECSN6 every 12 h starting from 2 h post-RV infection. A–C) TER was measured prior to RV infection (0 h) and at 24 h interval for up to 72 h during placebo or ECSN6 treatment. The data represent % change from TER at 0 h ± SEM from 3 experiments using cells from 3 donors (n = 3–12, cells from 3 donors). D) Apical surface of the cultures was washed 24 h post-infection, FITC-labeled inulin was added to the apical surface and fluorescence intensity was measured in the basolateral medium after 4 h. Data represents mean ± SEM calculated from cells obtained from 2 donors conducted in triplicates (n = 6). Statistical significance was conducted by ANOVA with Student-Newman-Keuls post-hoc analysis. * p < 0.05; ** p < 0.01; *** p < 0.001; ns = non-significant
Next, we examined whether reduction in TER corresponds to permeability, by assessing the paracellular movement of inulin 24 h post-RV-A1 infection. We chose 24 h post-infection since the reduction in RV-A1-infected cultures was maximum at this time. RV-A1-infected/placebo-treated cultures showed increased permeability to inulin coinciding with reduced TER (Fig. 1D). Treatment with 20% ECSN6 significantly reduced RV-A1-induced permeability to inulin. Together these results demonstrate that ECSN6 treatment prevents barrier disruption caused by RV-A1 infection.
ECSN6 prevents RV-A1-induced dissociation of E-cadherin and ZO-1Immunofluorescence microscopy was conducted to examine whether RV-A1 infection causes reduction in TER by dissociation of E-cadherin and ZO-1 as previously observed [7, 10]. RV-A1 caused dissociation of ZO-1 and E-cadherin from the intercellular junctions at 24 h post-infection and this was not observed in 20% ECSN6-treated cultures (Fig. 2). At 72 h post-infection, both ZO-1 and E-cadherin localization in RV-A1-infected/placebo-treated cultures returned to baseline (data not shown).
Fig. 2
ECSN6 inhibits RV-A1-induced dissociation of E-cadherin and ZO-1. Mucociliary-differentiated airway epithelial cultures were infected apically with RV-A1 or sham. The cultures were treated with placebo or 20% ECSN6 every 12 h starting from 2 h post-RV infection. At 24 h post-infection, cultures were fixed in cold methanol, blocked with 1% BSA and incubated with antibodies to ZO-1 and E-cadherin. The bound antibodies were detected by Alexa Fluor 488-labeled anti-mouse IgG (ZO-1) and Alexa Fluor 594-labeled anti-rabbit IgG (E-cadherin). Nuclei were counterstained with DAPI and imaged using confocal microscopy. Arrows in RV-A1-infected placebo-treated cultures represent dissociation of ZO-1 and E-cadherin. Images are representative of 3 experiments
To determine whether ECSN6 prevents RV-A1-induced reduction in TER by increasing the expression of adherence and tight junction molecules, we examined the protein expression of E-cadherin and a tight junction protein occludin by Western blot analysis using total proteins (Supplemental Fig. 3A). The ZO-1 antibody was not suitable for Western blot analysis, therefore we could not determine the expression of ZO-1. Quantification of band intensities demonstrated that RV-A1 increases E-cadherin at 72 h post-RV-A1 infection in both placebo and ECSN6 treated groups (Supplemental Fig. 3B). RV-A1-infection increased occludin protein expression in placebo-treated cells at 24 h post-infection, which returned to basal level after 72 h. Compared to RV-A1-infected placebo-treated cells, RV-A1-infected ECSN6-treated cells showed no increase in occludin at 24 h post-infection (Supplemental Fig. 3C), probably because RV-A1 did not disrupt intercellular junctions in ECSN6-treated cultures. Taken together, these results demonstrate that airway epithelial cells show increased expression of tight and adherence junction proteins in response to RV-A1-induced disruption of tight and adherence junctions probably to initiate repair processes. ECSN6 treatment did not affect the protein expression of adherence junction protein E-cadherin and tight junction protein occludin. These results indicate that ECSN6 may prevent RV-A1-induced reduction in TER by maintaining or enhancing the localization of E-cadherin and ZO-1 to the intercellular junctions and not by increasing the protein expression of E-cadherin or occludin.
ECSN6 reduces RV-A1-induced IL-6 and IL-8Since disruption of intercellular junctions can induce pro-inflammatory cytokine and chemokine expression, we determined whether ECSN6 reduces the expression of RV-A1-induced IL-8, IL-6, and CCL-20 at 24 h post-RV infection. As observed previously, RV infection caused significant increase in IL-8, IL-6 and CCL-20 [13] (Fig. 3A-C). Compared to placebo, 20% ECSN6-treated cell cultures showed reduction in RV-A1-induced IL-8 and IL-6, but not CCL-20. These results indicate that RV-A1-induced IL-6 and IL-8 may partially depend on disruption of intercellular junctions. ECSN6 maintains the integrity of intercellular junctions during RV-A1-infection, which in turn may lead to decreased RV-A1-induced pro-inflammatory responses.
Fig. 3
ECSN6 reduces RV-A1-induced pro-inflammatory IL-6 and IL-8 cytokine secretion. Mucociliary-differentiated airway epithelial cultures were infected apically with RV-A1 or sham. The cultures were treated with placebo or 20% ECSN6 every 12 h starting from 2 h post-RV infection. Basolateral medium was collected at 24 h post-infection. IL-6, IL-8 and CCL-20 protein levels were determined by ELISA. Since there was a wide variability from donor to donor in IL-8 levels in uninfected cells (sham/placebo) the data were normalized to sham/placebo. For IL-6 and CCL-20 the basal levels did not vary widely, therefore absolute levels are presented. Data represent mean ± SEM calculated from cells obtained from 3 donors in triplicates or more (n = 8–9). ANOVA with Student-Newman-Keuls post-hoc analysis was conducted to determine the statistical significance. * p < 0.05; ** p < 0.01; *** p < 0.001; ns = non-significant
ECSN6 increases the expression of antiviral mediators, but does not affect viral loadType I and type III IFNs are antiviral cytokines and expressed in response to viral replication [15, 16]. RV-A1 significantly induced IFN-β mRNA expression and IFN-λ1 and IFN-λ2 protein levels (Supplemental Fig. 4A-4C) in placebo-treated cells compared to sham. Please note that IFN-β protein levels were below detection levels, therefore we assessed mRNA levels. There was no difference in the mRNA expression of IFN-β between placebo and ECSN6 groups. However, RV-A1-induced IFN-λ1 and IFN-λ2 protein levels were slightly but significantly increased in 20% ECSN6-treated cells compared to placebo-treated cells. We also measured CXCL-10, another viral replication-dependent protein. RV-A1 infection significantly induced CXCL-10, which was slightly but significantly higher in ECSN6-treated cells (Supplemental Fig. 4D).
Since ECSN6 increased antiviral mediators, we examined the vRNA in the apical wash and in the cells. Interestingly, there was no difference in the vRNA between placebo and ECSN6-treated cultures in both extracellular and intracellular compartments at 24 h post-RV infection (Supplemental Fig. 4E and 4F).
ECSN6 prevents RV-A1-induced MUC5AC expressionAirway epithelial cells express both MUC5B and MUC5AC, however compared to MUC5B, MUC5AC is expressed at lower levels. As reported earlier, RV-A1 induced MUC5AC mRNA expression at 24 h post-infection [31]. Compared to placebo, ECSN6 significantly inhibited the mRNA levels of MUC5AC in RV-A1-infected cells (Fig. 4A). On the other hand, RV-A1 did not induce MUC5B mRNA expression (Fig. 4B).
Fig. 4
ECSN6 inhibits RV-A1-stimulated MUC5AC expression. Mucociliary-differentiated airway epithelial cultures were infected apically with RV-A1 or sham. The cultures were treated with placebo or 20% ECSN6 at 2 and 12 h post-RV infection. At 24 h post-infection, total RNA extracted from the cells was converted to cDNA and mRNA expression of MUC5AC, MUC5B and GAPDH was determined by qPCR. The expression of genes was normalized to GAPDH. Data represent mean ± SEM calculated from cells obtained from 3 donors in one to two replicates (n = 5). ANOVA with Student-Newman-Keuls post-hoc analysis was conducted to determine the statistical significance. * p < 0.05; ** p < 0.01; *** p < 0.001; ns = non-significant. ANOVA with Student-Newman-Keuls post-hoc analysis showed no difference between groups of MUC5B expression
Fig. 5
ECSN6 prevents RV-A1-induced reduction in CBF. Mucociliary-differentiated airway epithelial cultures were infected apically with RV-A1 or sham and treated with placebo or 20% ECSN6 every 12 h starting from 2 h post-RV infection. (A) At 24 h post-infection, the cultures were subjected to high-speed microscopy and CBF was analyzed by Ciliarmove. Data represent mean ± SEM calculated from cells obtained from 3 donors with duplicates or triplicates (n = 6–8). ANOVA with Student-Newman-Keuls post-hoc analysis was conducted to determine the statistical significance. * p < 0.05; ** p < 0.01; *** p < 0.001; ns = non-significant. (B) Cell cultures were fixed in paraformaldehyde, blocked with 1% BSA and incubated with antibody to acetylated α-tubulin. The bound antibody was detected by Alexa Fluor 488-labeled anti-mouse IgG. Nuclei were counterstained with DAPI and imaged using confocal microscopy. Representative images of cell cultures at 24 h post-RV-A1 infection. Yellow arrows in RV-A1-infected placebo-treated culture represent clumping of cilia
ECSN6 improves CBF in RV-A1-infected cellsExcessive mucus secretion, which occurs in response to viral infection, can transiently reduce CBF. Maintenance of cell polarity also plays an important role in regulating CBF. Since ECSN6 prevents dissociation of E-cadherin from adherence junctions to maintain the polarity of cells and inhibits MUC5AC expression, we postulated that ECSN6 may improve CBF. We measured CBF at 24 h, 48 h and 72 h post-RV-A1 infection and observed that RV-A1 attenuated CBF at 24 h (Fig. 5A), which returned to baseline by 48 h (data not shown), indicating that the virus transiently reduces CBF. Interestingly, the transient reduction in CBF by RV-A1 was inhibited by 20% ECSN6. Qualitatively, there was no difference in the number of ciliated cells (Fig. 5B). While cilia on the cells appeared well-separated in sham-infected, placebo or ECSN6-treated cultures, cilia seemed to be clumped in RV-A1-infected placebo-treated cells. Such clumping of cilia was not observed in RV-A1-infected and ECSN6-treated cells and cilia look similar to that observed in sham-infected cultures.
Combination of prophylactic and therapeutic treatment with ECSN6 protects mucosal barrier function during RV-A1 infectionNext, we examined whether pretreatment with ECSN6 (prophylactic) combined with post-RV-A1 infection treatment (therapeutic) is better or as good as therapeutic strategy in attenuating RV-A1-induced disruption of TER, IL-8 release and CBF attenuation. Cell cultures were pretreated with placebo or 20% ECSN6 for 48 h, infected with RV-A1 and treated with placebo or 20% ECSN6 twice a day for another 24 h. As observed earlier, RV-A1 infection reduced TER and CBF in placebo-treated cultures (Supplemental Fig. 5A and 5C). These RV-A1-induced changes were significantly inhibited by ECSN6 treatment. In both placebo- and ECSN6-treated cultures, RV-A1 increased IL-8, but it was significantly lower in the latter group (Supplemental Fig. 5B). As observed in cultures treated after RV infection, there was no difference in the vRNA between prophylactically placebo- and ECSN6-treated cultures in intracellular compartments at 24 h (Supplemental Fig. 5D). In addition, immunolocalization studies showed that ECSN6 treatment inhibits RV-induced dissociation of both ZO-1 and E-cadherin from the intercellular junctions (Supplemental Fig. 5E). Taken together, these results demonstrate that combination of prophylactic and therapeutic treatment strategy is as effective as therapeutic strategy in protecting mucosal barrier function during RV-A1 infection in airway epithelial cell cultures.
ECSN6 protects nasal epithelial cells from RV-A1-induced pathologic changesSince nasal epithelium is the primary target for RV, we investigated whether ECSN6 protects the nasal epithelium in a similar way to tracheal epithelium from RV-induced pathologic changes using nasal cells from two healthy non-smokers. Treatment with 20% ECSN6 increased TER in sham-infected cell cultures (Supplemental Fig. 6A). Compared to sham-infected and placebo-treated cell cultures, mucociliary-differentiated nasal epithelial cell cultures infected with RV-A1 and treated with placebo showed reduced TER and CBF, and increased the expression of IL-8 (Supplemental Fig. 6A to 6C). Cell cultures treated with ECSN6 prevented RV-A1-induced reduction in TER and CBF. Treatment with ECSN6 also reduced RV-A1-induced IL-8 release. These results indicate that ECSN6 treatment also improves barrier function in nasal epithelial cell cultures, thus reducing pro-inflammatory responses and improving CBF.
Treatment with ECSN6 protects mucosal barrier function during RV16 infectionRV16 belongs to major group RV and binds to human ICAM-1. Previously, we have demonstrated that RV39 which is also a major group RV, disrupts barrier function similar to RV-A1 [7]. Here, we examined whether ECSN6 also prevents the effects of human ICAM-1 binding RV16-induced pathologic changes using mucociliary-differentiated tracheal epithelial cell cultures. Similar to RV-A1, RV16 significantly reduced TER, increased the permeability to inulin, and caused dissociation of E-cadherin and ZO-1 at 24 h post-infection. ECSN6 completely inhibited these RV16-induced pathological changes (Supplemental Fig. 7A – 7D). As observed with RV-A1, at 72 h post-RV16 infection, TER returned to basal levels in placebo-treated cultures and even increased in ECSN6-treated cultures when compared to the respective placebo group. Western blot analysis showed no difference in the expression of E-cadherin and a tight junction protein occludin between placebo and ECSN6 treated cultures (data not shown) indicating that ECSN6 may prevent RV16-induced reduction in TER by maintaining or enhancing the localization of E-cadherin and ZO-1 to the intercellular junctions.
ECSN6 significantly reduced RV16-induced IL-6, but not CCL-20 and IL-8 at 24 h post-infection (Supplemental Fig. 8A –8C). Unlike RV-A1, RV16 induced both, MUC5B and MUC5AC at 24 h post-infection and this was inhibited by ECSN6 (Supplemental Fig. 8D and 8E). We also determined the antiviral IFN responses and viral RNA copy number. RV16-infected cell cultures show significantly higher mRNA expression of both IFN-β and IFN-ʎ1 than sham-infected cultures. Compared to RV16-infected placebo treated cultures, similarly infected ECSN6-treated cultures showed small but significant increases in the expression of antiviral molecules, IFN-β and IFN-ʎ1 (Supplemental Fig. 8F and 8G). Again, ECSN6 had no effect on viral load (data not shown).
Taken together these results suggest that ECSN6 treatment similarly affects RV-A1- and RV16-induced effects on barrier dysfunction, pro-inflammatory IL-6 responses, and mucin expression.
ECSN6 reduces inflammation and prevents dissociation of E-cadherin induced by RV-A1 in vivoIn the initial experiments, we examined the tolerance of ECSN6 in mice by treating mice intranasally with 20, 40, or 60% ECSN6 for up to 60 h twice a day and then examined for nasal inflammation by histology and mRNA expression of pro-inflammatory cytokines 12 h after the last treatment. Mice treated with 20 and 40% drug showed normal histology and were similar to mice treated with PBS (control group) (Supplemental Fig. 9). Mice treated with 60% ECSN6 showed infiltration of inflammatory cells (Supplemental Fig. 9E, arrows) and significant increase in the mRNA expression of some pro-inflammatory cytokines, such as Cxcl2, Tnf-α, and Il-6 (Supplemental Fig. 10). Therefore, we used 40% ECSN6 which did not cause inflammation in the subsequent experiments.
Mice infected intranasally with RV-A1 or sham were treated with placebo or 40% ECSN6 twice a day at 10 h interval starting from 1 h after the infection for up to 60 h and examined for histological changes in the nose at 24 h post-infection. The expression of pro-inflammatory cytokines, chemokines and mucin genes were determined at 24 h and 72 h post-infection using TRIZOL lysates of sinunasal mucosa. At 24 h post-infection, sham-infected mice did not show changes in histology or mucus secretion irrespective of the treatment. Compared to sham-infected animals, mice infected with RV-A1 and treated with placebo showed increased mucus secretion with patchy inflammation in the sinunasal cavities (Fig. 6A and B). RV-A1-infected mice treated with ECSN6 showed less inflammation and reduced mucus secretion compared to placebo-treated mice. To confirm these findings, the inflammation at different depths of the snout was scored by a veterinary pathologist who was blinded for treatment and infection. The inflammatory scores included accumulation of secretions, infiltration of mononuclear cells and neutrophils into the sinunasal cavities. Scoring of inflammatory changes indicated moderate inflammation in RV-A1-infected animals treated with placebo. In contrast similarly infected ECSN6-treated animals showed very mild to mild inflammation, which was significantly lower than in similarly infected placebo-treated animals (Supplemental Table 2). There was no difference in the number of goblet cells/100 µM between the groups (data not shown). Interestingly, in representative histopathology images, the cilia in some part of the sinunasal epithelium showed mild disorganization in the RV-A1-infected placebo-treated group, but not in other groups (Fig. 6C, arrows).
Fig. 6
RV-A1-infected mice treated with ECSN6 show reduced inflammation and mucus secretion. Mice were infected with sham or RV-A1 and treated with placebo or 40% ECSN6 twice a day at 10 h interval for 24 h starting from 1 h after sham infection. The paraffin sections of snout in (A and C) were stained with H & E or in (B) with PAS. * represents sinunasal cavity and inflammatory milieu respectively in panel A; red and yellow arrows represent secreted mucus and goblet cells, respectively in panel B; and black arrow represent disorganization of cilia in panel C. Images are representative of 3–4 mice in each group
Compared to sham, RV-A1 increased mRNA expression of Cxcl2, Tnf-α and Muc5ac at 24 h post-infection in placebo-treated groups. Compared to sham-infected and placebo-treated groups, RV-A1-infected and ECSN6-treated animals showed increase in Cxcl2 and Cxcl10 at 24 h post infection but not Tnf-α and Muc5ac. At 72 h post-infection, RV-A1-infected placebo-treated mice showed persistent expression of Cxcl2, Tnf- α and expression of Cxcl10, while ECSN6-treated mice showed persistent expression of Cxcl10 and no significant increase in the expression of Cxcl2 and Tnf-α compared to sham-infected and placebo-treated mice (Fig. 7A and E).
Fig. 7
ECSN6 attenuates RV-A1-induced expression of pro-inflammatory cytokines. Mice were infected with sham or RV-A1 and treated with placebo or 40% ECSN6 twice day at 10 h interval for up to 60 h starting from 1 h after RV-A1 infection. Mice were euthanized after 24–72 h post-infection and total RNA from sinunasal TRIZOL lysates was isolated and subjected RT-qPCR using gene-specific Taqman assays. The mRNA expression was normalized to β-actin. Data represent median with range from 3 experiments with a total of 6 mice per group (n = 6). ANOVA on ranks with Tukey post-hoc analysis. * p < 0.05; ** p < 0.01; *** p < 0.001; ns = non-significant. ANOVA on ranks with Tukey post-hoc analysis showed no difference between groups of Muc5B expression at 24 h and 72 h post-infection and Muc5ac expression at 72 h post-infection
Interestingly, mice infected with RV-A1 and treated with placebo showed dissociation of E-cadherin from the intercellular junction of nasal epithelium, and this was prevented by treatment with ECSN6 (Fig. 8A). Analysis of E-cadherin density by Image J indicated that compared to sham, RV-A1-infected animals show significant reduction in E-cadherin protein density in placebo treated group but not in ECSN6 group (Fig. 8B). However, there was no difference in the mRNA expression of E-cadherin irrespective of infection or treatment (Fig. 8C). These results may suggest that ECSN6 may prevent RV-A1-induced dissociation of E-cadherin by maintaining or enhancing the localization of E-cadherin and not by increasing the expression of E-cadherin.
Fig. 8
ECSN6 treatment prevents RV-A1-induced dissociation of E-cadherin. Mice were infected with sham or RV-A1 and treated with placebo or 40% ECSN6 twice day at 10 h interval for up to 60 h starting from 1 h after RV-A1 infection. (A) At 24 h post-infection, the paraffin sections of snout were prepared and immunostained with E-cadherin and imaged under a fluorescence microscope. Arrow in RV-A1-infected placebo-treated panel denotes dissociation of E-cadherin from the intercellular junctions of nasal epithelium. Images are representative of 3 to 4 mice per group. (B) Density of E-cadherin was measured by Image J and expressed as pixels/50 µm2. Data represent median with range and statistical significance was determined by ANOVA on ranks with Tukey post-hoc analysis (n = 3 to 4; *p = < 0.05). (C) Total RNA from sinunasal TRIZOL lysates was isolated and subjected RT-qPCR using gene-specific Taqman assays after 24–72 h post-infection. The mRNA expression was normalized to β-actin. Data represent median with range from 3 experiments with a total of 6 mice per group. ANOVA on ranks with Tukey post-hoc analysis showed no difference between groups
Treatment with ECSN6 had no effect on RV-induced mRNA expression of the antiviral genes Ifn-λ1 and Ifn-β nor reduced the viral load in mice infected with RV-A1 as observed in airway epithelial cell cultures (data not shown).
Taken together, these results demonstrate that ECSN6 prevents RV-A1-induced dissociation of E-cadherin from intercellular junctions of sinunasal epithelium, reduces inflammation and accumulation of secretions in sinunasal cavities, and appears to prevent cilia disorganization of murine airway epithelium even though no effect on viral clearance was observed. In addition, the reduced inflammation and mucus secretion was associated with a beneficial influence on RV-A1-induced cytokine responses and Muc5ac mRNA expression.
留言 (0)