Tumor stem cells represent a heterogeneous and plastic subpopulation of cells, capable of both self-renewal and generation of progenitor cells that will be subject to the differentiation process [18]. Furthermore, these cells contribute to resistance to conventional therapies and to hostile microenvironmental conditions, playing a role in disease recurrence [19]. Therefore, studies that describe efficient strategies to identify these cells are of great value, as they allow a better characterization and the development of new therapeutic approaches that could target tumor stem cells. In this context, the present study aimed to identify and characterize glycobiomarkers present in the CSC population.
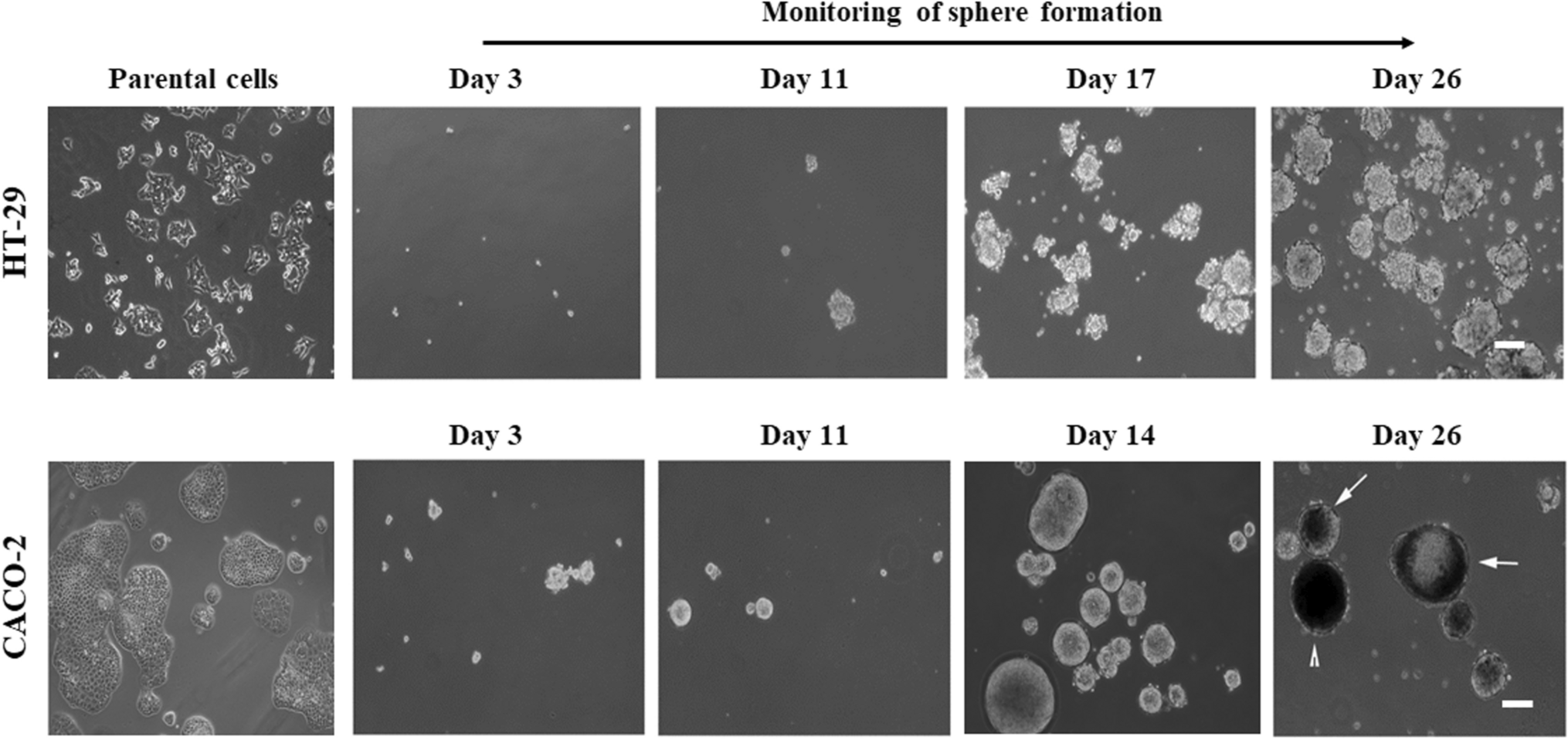
For this purpose, we employed the sphere formation assay, a well-established method for enriching cancer stem cells (CSCs). In this assay, cells at clonal density (the density that allows clonal generation of spheres) are challenged to grow under non-adherent conditions in a medium that promotes the enrichment of CSCs and lacks differentiation inducers. This setup mimics the behavior of CSCs, which are known to survive and proliferate in such non-adherent, serum-free conditions, reflecting their self-renewal and undifferentiated properties [20, 21]. As such, the formation of spheres is a functional indicator of the presence of CSCs within a population. The success of this assay depends on critical parameters such as medium composition, cell density, and exposure time to the medium [22]. In this study, we standardized a sphere formation assay protocol for the HT-29 and Caco-2 colon adenocarcinoma cell lines. Both cell lines successfully formed spheres, but with different shapes, sizes, and timeframes. This variation could reflect an experimental limitation, which may introduce greater variability when comparing independent experiments. Routinely, the medium used is DMEM/F12 supplemented with EGF, FGF, B27, insulin, hydrocortisone, BSA, and heparin, without the addition of FBS [23,24,25]. However, as cells are plated from a 2D culture, in which not all cells have stem cell capacity, there is considerable experimental variability, with spheres varying in size. Additionally, the amount of material generated by this assay is often small, limiting the feasibility of performing analyses that require large numbers of cells.
Once the sphere formation protocol was standardized, it was necessary to ensure that the obtained population was enriched for CSCs. For this, the expression of the CSC markers CD44, CD24, PROM1, ALDH1 and LGR5 was evaluated by RT-qPCR. When comparing the sphere condition with the parental condition, a significant increase in the expression of ALDH1, LGR5, and PROM1 was observed in HT-29, while in CACO-2, a significant reduction in the expression of CD24 was detected. Although both cell lines were incubated in the same CSC enrichment medium, the observed expression differences highlight the heterogeneity of the CSC population. A similar heterogeneity was portrayed by a previous study analyzing five CRC lineages, including HT-29 and CACO-2. In this case, the CD133+ population of each of the analyzed cell lines was isolated and other markers related to stemness, tumorigenesis and metastatic potential were evaluated. In conclusion, the study reported that three sets of markers were found: those that were detected in all cell lines; those that presented heterogeneous detection, that is, differences in the percentage of cells expressing them; and those markers with discontinuous marking, sometimes present and sometimes absent, depending on the cellular model [26]. Therefore, the differences observed in the expression profile of CSCs markers in HT-29 and CACO-2 may reflect the presence of subpopulations with distinct profiles in the spheroids.
At first, the reduction in CD24 expression in CACO-2 seems contradictory, since the increase in this marker has already been documented in CSCs populations from CRC lines [27]. However, this reduction could indicate the presence of a CSCs subpopulation that present mechanisms of drug resistance and survival mediated by autophagy. Since the silencing of CD24 in cells that overexpressed this marker promoted an increase in autophagy in CRC cells [28]. In this sense, it was also demonstrated that the inhibition of the autophagic process was capable of sensitizing CRC cells to 5-Fluorouracil (5-FU) and antiangiogenic drugs [29, 30]. Corroborating these findings, it was described in CRC that the CD133+/CD24low phenotype is associated with resistance to 5-FU [31].
From the oncogenic viewpoint, the expression of certain glycan structures provides advantages in the malignant transformation, since these carbohydrates are involved in processes such as: migration, invasion, angiogenesis, immune surveillance, and metastasis [32, 33]. In addition, considering that many of the CSCs markers are in fact glycoproteins we thought it would be interesting to investigate whether any of the glycogenes altered in the sphere formation assay could be correlated to their expression and, consequently impact any cellular mechanisms in which they are involved.
Thereby the sphere formation assay results showed an enhancement in the expression of B3GNT2, OGT and OGA in the HT-29 spheres related to the parental cells. Furthermore, to our knowledge, this is the first time in the cancer glycobiology field, that in silico analyzes pointed to the existence of a positive correlation between the expression of a CSC marker, LGR5, and a glycogene, OGA, in rectal and colon cancer. In both cases, the correlations were significant, however, weak (r between 0.20 and 0.39) or very weak (r between 0.00 and 0.19) [34]. Due to the low frequency of CSCs in the tumor masses, it can be difficult to detect their expression in tumor samples, weakening any correlations involving them. It is worth mentioning that the positive correlation between the expression of LGR5 and OGA suggests a possible regulation between these two genes, since, as demonstrated in the present study, the sphere formation assay promotes an increase in the expression of both in HT-29. Although it can be speculated that this glycogene may influence the stemness and the expression of CSCs markers in colorectal cancer, the potential mechanisms involved are not yet known, showing that future studies will be necessary to clarify this aspect.
Another relevant observation was the increased expression of OGT and OGA in spheroids. The OGT, also known as the O-GlcNAc transferase, is the enzyme responsible for the incorporation of a GlcNAc motif to the Ser/Thr of proteins in a post-translational process called O-GlcNAcylation. Alternatively, O-GlcNAcylated proteins can be reverted to its deglycosylated state due to the removal of this motif, which is catalyzed by the OGA enzyme, also known as O-GlcNAcase [35]. Bearing this in mind, it was recently demonstrated that in spheres from breast cancer lines there is a concomitant increase in OGT and O-GlcNAcylation. Furthermore, OGT inhibition had a negative impact on the stemness of these cells. Its overexpression had the opposite effect, leading to the upregulation of CD44, NANOG, MYC and KLF8, indicating that the increase in OGT levels in the spheres may be necessary for the regulation of transcriptional factors related to pluripotency [36].
As the presence of stemness characteristics in circulating tumor cells has been observed before in patients with metastatic CRC [37], this finding motivated the evaluation of the clinical impact of the changes in the expression of CSCs markers and glycogenes in patients with CRC. Although we did not observe a significant difference in the expression of the CSC signature (ALDH1, LGR5 and PROM1) between normal and tumor samples, in rectal cancer specimens, LGR5 expression was increased in all tumor stages relative to normal tissue. On the other hand, the increase in the expression of PROM1 was only observed in the stages I, II, and III. At the same time, a reduction in ALDH1 expression could be detected in all stages. In colon carcinoma samples, the same scenario was observed for LGR5 expression, while for ALDH1 a reduction from stage II onwards was observed. PROM1 had significant differences in its expression from the stages I and II relative to stage IV. However, only the ALDH1 expression could predict the overall survival of rectal carcinoma patients (representing a 5.5-fold increased risk of death for the group with a higher expression), while none of the other markers analyzed was associated with disease-free survival or overall survival. This lack of association could be related to the fact that the analyses were based on gene expression and not protein levels.
The data found in silico by us is corroborated at least in part by a previous study that showed by immunohistochemistry a slight ALDH1 reduction in colorectal tumors in comparison with normal samples [38]. Another study also corroborates the findings by pointing out increased LGR5 labeling in CRC tissue samples [39].
The same in silico analyses conducted for the CSC signature were also applied to glycogenes. However, no differences were observed in the expression of the glycogene signature (OGT, OGA and B3GNT2) between normal and tumor samples. However, OGT expression was increased throughout the progression of rectal and colon carcinoma. In the latter, OGT expression levels increased from stage II to IV, which might suggest a role for OGT in the establishment of distant metastases. In fact, higher OGT expression was positively associated with the total staging, and the nodal and metastasis stages before [40]. Corroborating the literature associations, our study found a glycogene signature capable of predicting the overall survival for the colon carcinoma cohort. Moreover, when these analyses were conducted individually the higher OGA expression alone was associated with an increased risk of recurrence in rectal carcinoma, and, also with a higher risk of death for the colon carcinoma patients. Interestingly, a previous study described that Apcmin/+Oga± mice had less and smaller colorectal adenomas when compared to the Apcmin/+ mice, due to a reduction in the expression of target genes involved in the WNT pathway in a β-catenin independent manner [41]. Therefore, the augment in the OGA expression observed in the sphere condition may occur to induce the expression of WNT target genes. It is important to highlight that this pathway is related to the CSCs maintenance [42]. In addition, the LGR5 expression can be modulated by the WNT signaling pathway as it represents one of its target genes [43]. This substantiates the positive correlation found between the LGR5 and OGA expressions and indicates a conceivable mechanism to explain the OGA effect on patients’ reduced survival. Other hypothesis that could explain the results seen in the patients’ survival is that OGA could be impairing the p53 expression by removing the PDLIM7 O-GlcNAcylation. As it was recently described, the removal of the PDLIM7 O-GlcNAcylation by OGA affects the p53 expression inducing its proteasome degradation and diminishing its transcription [44]. However, further investigations are needed to confirm these inferences.
It is important to highlight that all changes observed in the present study were investigated in relation to gene expression and do not necessarily reflect changes at the protein level or even reflect greater or lesser presence of a particular glycan arrangement. Therefore, complementary analyzes of the protein levels of CSC markers, the enzymes encoded by glycogenes, the arrangements generated by them, as well as the proper signaling pathways leading to the alterations observed in our study need to be addressed in further investigations.




留言 (0)