
記住我
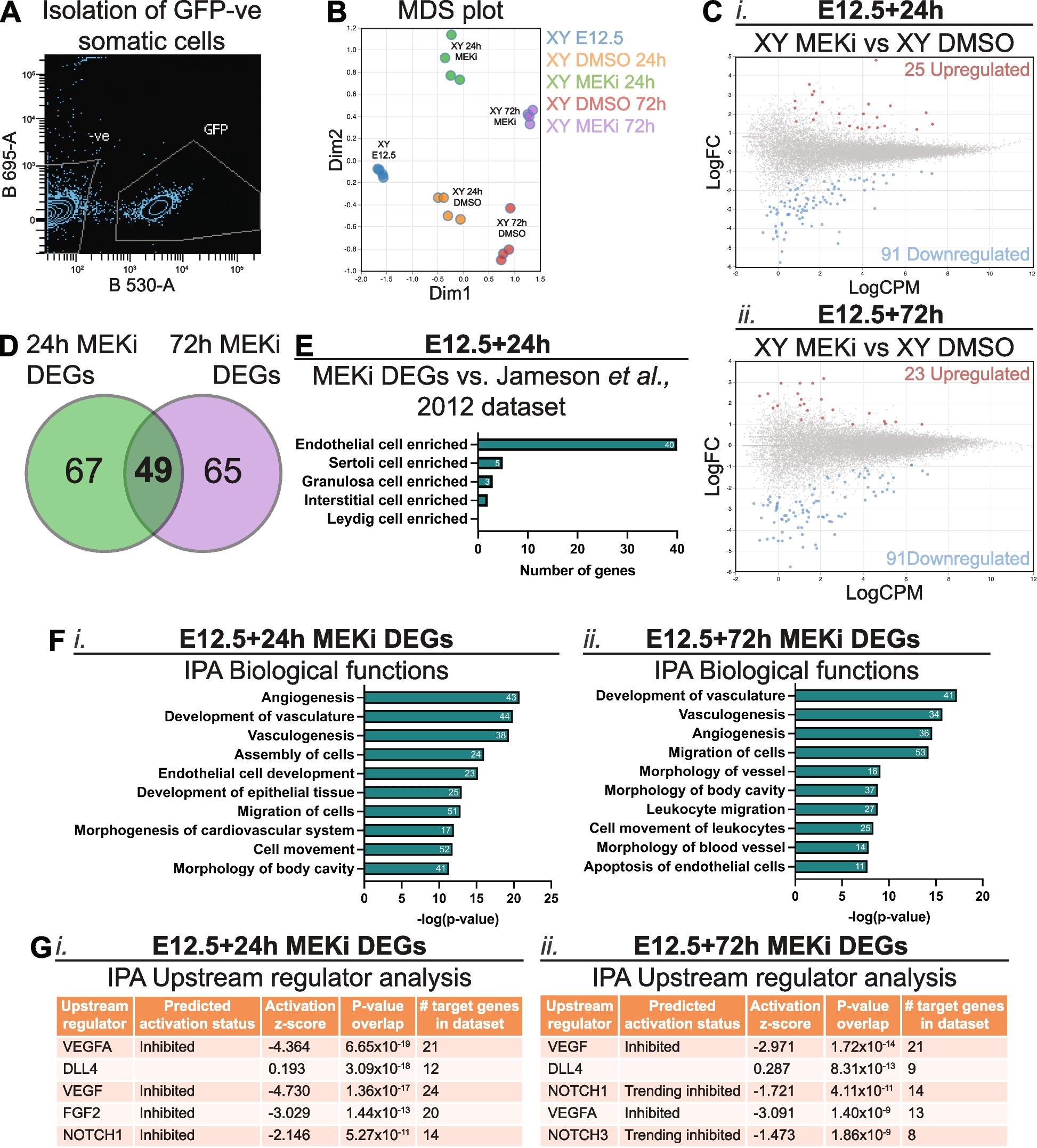





We previously demonstrated that MEK1/2 inhibition disrupts Sertoli cell proliferation and organisation [13]. To investigate the role of MEK1/2 signalling in the somatic cells of developing testis further, Oct4 (Pou5f1)-eGFP (OG2) transgenic male mice (129T2svJ background) [39,40,41] were crossed to Swiss females and E12.5 testis-mesonephros samples were isolated and cultured for 24 or 72h with DMSO or the small molecule inhibitor PD0325901/Mirdametinib (MEKi) at a concentration of 500nM. This dose was selected based on previous titration experiments that identified a minimal dose that functionally altered germ cell mitotic arrest and Sertoli cell proliferation, and completely abrogated the ability of FGF9 to promote somatic cell proliferation in E12.5 XX gonads [13]. To gain insight into genome-wide transcriptional changes that occur in the somatic cells of MEK1/2 inhibited testes, RNA sequencing was performed on testicular somatic cells obtained by fluorescent activated cell sorting (FACS) which isolated Oct4-GFP negative cells of E12.5 XY gonads cultured for 24 or 72h (Fig. 1A; [40, 41]. The RNA sequencing data from each treatment group clustered independently in a multidimensional scaling plot (Fig. 1B), indicating that inhibition of MEK1/2 signalling altered somatic cell transcription. Further analysis identified 116 and 114 differentially expressed genes (DEGs) following culture in MEKi for 24 or 72h (Additional file 1: Table S1.1 and S1.2), respectively (false discovery rate (FDR) < 0.05, absolute fold-change (FC) ≥ 1.5, absolute log-FC ≥ 0.585; Fig. 1C). Comparison of the DEGs identified 49 genes which were commonly disrupted after 24 or 72h of MEKi (Fig. 1D; Additional file 1: Table S1.3). These included endothelial associated genes such as Pecam1 (Cd31), Flt1 (Vegfr1), Ecscr, and Tie1, the angiogenesis marker Sox7, and the cell-surface glycoprotein encoding gene Cd93, which has been associated with enhanced macrophage phagocytosis and inter-cellular adhesion [42]. In addition, 67 genes were uniquely disrupted after 24h of MEKi (Fig. 1D; Additional file 1: Table S1.3) including the gap junction genes Gja4 and Gja5, the angiogenesis marker Sox17, and the chemokine receptor Cxcr4. Similarly, MEK1/2 inhibition for 72h exclusively disrupted 65 genes (Fig. 1D; Additional file 1: Table S1.3), including the retinoic acid degrading enzyme coding gene Cyp26b1 and the type IV collagen coding genes Col4a1 and Col4a2, which function downstream of Wt1 to support testis cord integrity in the fetal mouse testis [43].
Fig. 1
MEK1/2 signalling is required for expression of genes associated with endothelial cells, vasculogenesis and angiogenesis in somatic cells of developing testes. RNA sequencing analysis of somatic cells isolated from E12.5 testes and E12.5 testes cultured for 24 or 72h with DMSO or 500nM of MEKi. A Example of FACS scatterplot depicting isolation of GFP negative somatic cells. B Multidimensional scaling (MDS) plot of somatic cell samples subject to RNA sequencing. C Differential gene expression analysis of E12.5 + 24 h (i) or E12.5 + 72 h (ii) MEKi vs DMSO. Genes with FDR < 0.05 and |logFC|≥ 0.585 (equivalent to |FC|≥ 1.5) were considered differentially expressed. D Venn Diagram comparing somatic cell RNA sequencing data from E12.5 testis samples cultured with DMSO or MEKi 24h and 72h to identify common differentially expressed genes (DEGs). E Comparison of 24h MEKi DEGs with the microarray data set published by Jameson et al., [44], PLOS Genetics. F Ingenuity Pathway Analysis (IPA) of biological functions associated with 24h (i) and 72h (ii) MEKi DEGs. The number of genes associated with each term is shown within the bar. G IPA of upstream regulators associated with 24h (i) and 72h (ii) MEKi DEGs. Biological replicates; n = 4 per treatment
The somatic cell population of the fetal testis and ovary is comprised of supporting cells (Sertoli – XY and granulosa – XX) which surround germ cells in the XY fetal gonad, stromal (XX) and interstitial (XY) cells which give rise to steroidogenic theca (XX) and Leydig (XY) cells, endothelial cells, which contribute to vasculature and cord formation in XY gonads, and immune cells [44]. To determine if particular cell populations were affected by MEKi more than others, we compared our MEKi DEGs with published data [44]. As Jameson et al., [44] assessed E11.5–13.5 gonads and our samples are most closely aligned in age with E13.5 gonads, we compared our 24h MEKi DEGs with the E13.5 data. Of the 116 DEGs identified, 40 were specifically enriched in endothelial cells, five in Sertoli cells, three in granulosa cells and two in interstitial cells at E13.5 (Fig. 1E, Additional file 1: Table S1.4). All 40 genes specifically enriched in endothelial cells were down regulated after 24h MEKi treatment indicating either that endothelial cells are lost, that transcription of these genes depends on MEK1/2 signalling or both (Additional file 1: Table S1.4). The remaining genes were not specifically enriched in any cell type (46 genes) or were not detected in the microarray data set (20 genes; Additional file 1: Table S1.4).
To determine the biological functions affected by MEKi, we assessed the 24h and 72h MEKi DEGs using Ingenuity Pathway Analysis (IPA; Fig. 1F). IPA determined that many 24 h MEKi DEGs were associated with angiogenesis (43 genes), development of vasculature (44 genes), and vasculogenesis (38 genes), assembly of cells (24 genes) and migration of cells (51 genes; Fig. 1Fi). Some of these terms were also associated with MEKi 72h DEGs including, development of vasculature (41 genes), vasculogenesis (34 genes), angiogenesis (36 genes), and migration of cells (53 genes; Fig. 1Fii). Other terms including morphology of vessel (16 genes) and apoptosis of endothelial cells (11 genes) were also associated with MEKi 72h DEGs. Although terms relating to cell death or apoptosis were not present in the top 10 most associated biological functions in the 24h MEKi DEGs, cell death of endothelial cells was the 27th associated term, while apoptosis was 29th and apoptosis of endothelial cells was 31st. Moreover, IPA upstream regulator analysis predicted upstream pathways associated with the 24h MEKi DEGs including VEGFA, DLL4, VEGF, FGF2 and NOTCH1 (Fig. 1Gi). Unsurprisingly, the top five predicted upstream regulators of the 72h MEKi DEGs were similar to the 24h MEKi DEGs and included VEGF, DLL4, NOTCH1, VEGFA and NOTCH3 (Fig. 1Gii). Together these analyses highlight very strong endothelial cell, angiogenesis and vasculogenesis signatures in testes following MEK1/2 inhibition, indicating that MEK1/2 may act downstream of VEGFA/VEGF signalling to promote the expansion and organisation of the fetal testis vasculature.
MEK1/2 signalling is essential for endothelial cell development in fetal testesTo determine if MEK1/2 signalling acts directly in endothelial cells, we used IF to assess the localisation of the downstream target of MEK1/2, pERK1/2 and the endothelial cell marker, CD31, in E12.5, E13.5, E14.5 and E15.5 testes (Additional file 2: Fig. S1). At E12.5, strong pERK1/2 staining was detected in CD31 positive endothelial cells but not in germ cells, which had weaker CD31 staining and were clearly morphologically distinguishable from endothelial cells. Although less intense than the staining observed in endothelial cells, pERK1/2 was also detected in cells immediately surrounding germ cells. Given their punctate DAPI staining (marking DNA) and the location of these somatic cells inside testis cords, we concluded that these were Sertoli cells. Intense pERK1/2 staining was maintained in endothelial cells in E13.5 testes, but pERK1/2 was no longer detected in Sertoli cells. At E14.5, pERK1/2 was detected in endothelial cells and was again detected in Sertoli cells. Furthermore, bright pERK1/2 staining was detected in Sertoli cells located in the middle of the testis cords, and less intense staining was detected in Sertoli cells towards the testis cord basement membrane. At E15.5, endothelial cells were pERK1/2 positive and the staining in Sertoli cells remained similar to that detected in E14.5 Sertoli cells.
To confirm that MEKi inhibited MEK1/2 activity, we examined pERK1/2 localisation in E12.5 testes cultured for 24h or 72h in DMSO or MEKi media. While pERK1/2 was detected in DMSO controls, no pERK1/2 was detected in testes following 24h or 72h MEKi treatment (Fig. 2, Additional file 3: Fig. S2, Additional files 4–7: Movies S1-4). In 24h DMSO controls, pERK1/2 was detected in the CD31 positive endothelial cells, with strong pERK1/2 staining in cells of the coelomic vessel (Fig. 2A, Additional file 3: Fig. S2Ai). Moreover, weaker pERK1/2 staining was detected in Sertoli cells located in testis cords and surrounding germ cells (Fig. 2A, Additional file 3: Fig. S2Ai). Co-staining pERK1/2 with SOX9 confirmed this observation, with pERK1/2 detected in the nucleus or cytoplasm of some Sertoli cells, but not detected in other Sertoli cells possibly depending on their cell cycle state (Additional file 3: Fig. S2B). Notably, while pERK1/2 positive Sertoli cells were detected in E12.5 testes cultured for 24h (Additional file 3: Fig. S2B), pERK1/2 was not detected in E13.5 in vivo collected testes, although it was detected at E12.5 and E14.5 (Additional file 2: Fig. S1). This difference in pERK1/2 detection in E13.5 in vivo and E12.5 + 24h cultured testes is presumably due to slightly delayed development caused by isolation of E12.5 gonads and setting them in culture, consistent with transient loss of pERK1/2 in Sertoli cells between E12.5 and E13.5 (Additional file 2: Fig. S1).
Fig. 2
MEK1/2 inhibition depletes pERK1/2 and CD31 expressing endothelial cells in developing testes. Section or wholemount immunofluorescent imaging of E12.5 testes cultured for 24 or 72h with DMSO or 500nM of MEKi. A and C Section IF of cultured testes stained with DAPI (blue), pERK1/2 (red) and CD31 (endothelial cells and germ cells; cyan). B Three-dimensional stacked Z-series images of cultured testes showing Oct4-eGFP (germ cells; green), CD31 (endothelial cells and germ cells; blue) and pERK1/2 (red). The DMSO and MEKi images should be viewed in conjunction with Additional files 4–7: Movies S1-4, which reveal the 3D organisation of the cells in the tissue. Scale bar represents 100 μm. Arrows indicate pERK1/2 positive endothelial cells and asterisks indicate pERK1/2 positive Sertoli cells. Biological replicates; n = 3–4 per treatment
To gain more detailed visualisation of endothelial cells in three-dimensional space, we cultured gonads for 24h with DMSO or MEKi in hanging drops and used wholemount IF to examine CD31 and pERK1/2. Wholemount IF revealed a similar pattern of Oct4-eGFP, pERK1/2 and CD31 in E12.5 testes cultured with DMSO, with pERK1/2 detected in CD31 positive endothelial cells particularly towards to coelomic epithelium, in between the testis cords and in the gonad-mesonephric border (Fig. 2B, Additional files 4–5: Movie S1-2). In addition, Oct4-eGFP negative Sertoli cells detected within the testis cords were pERK1/2 positive (Fig. 2B, Additional files 4–5: Movies S1-2). In contrast, no pERK1/2 positive cells were detected in 24h MEKi treated testes using section IF (Fig. 2A; Additional file 3: Fig. S2Ai). Furthermore, CD31 positive endothelial cells were not detected in these gonads indicating that vasculature was lost within the testis, although some CD31 positive endothelial cells were detected in the mesonephros (Fig. 2A, Additional file 3: Fig. S2A). This was confirmed with wholemount IFs of MEKi treated testes, with no pERK1/2 detected and the number of CD31 positive endothelial cells within the testes greatly reduced, although some endothelial cells were detected in the mesonephros (Fig. 2B, Additional files 6–7: Movies S3-4). Consistent with these findings, pERK1/2, CD31 double positive endothelial cells marked the coelomic vessel in 72h DMSO cultures (Fig. 2C, Additional file 3: S2Aii). In addition, pERK1/2 was detected in Sertoli cells, although the staining intensity was weaker than in endothelial cells (Fig. 2C, Additional file 3: Fig. S2Aii). In contrast, pERK1/2 was not detected, and CD31 positive endothelial cells were not observed in testes following 72h of culture with MEKi (Fig. 2C, Additional file 3: Fig. S2Aii). This was consistent with 3.99- and 5.94-fold reduced Cd31 transcription observed by RNA sequencing after 24 and 72h MEKi treatment respectively (Additional file 1: Table S1.1 and S1.2), confirming that MEKi reduced both transcription and protein levels of CD31. While it is possible that MEK1/2 inhibition resulted only in loss of CD31 expression and endothelial cells remained, RNA sequencing analysis revealed lower transcriptional levels of all 40 DEGs that were endothelial cell enriched genes, strongly indicating that endothelial cells were lost (Fig. 1E; Additional file 1: Table 1.4). Moreover, IF staining for the definitive endothelial marker VE-Cadherin specifically marked well-organised endothelial cells in DMSO controls after 24h of culture, but no VE-Cadherin positive cells remained within the gonad of MEKi-treated samples (Additional file 8: Fig. S3). This was consistent with 6.7-fold reduction in Cdh5 (encodes VE-cadherin) transcription after 24h MEK1/2 inhibition observed by RNA sequencing (Additional file 1: Table 1.1). Together these data indicate that inhibiting MEK1/2 signalling severely impacted vascular development in E12.5 testes.
MEK1/2 inhibition results in loss of the angiogenic transcription factors SOX7 and SOX17 in testicular endothelial cellsSox7 and Sox17 are genes in the SoxF subgroup that regulate angiogenesis and vasculogenesis [21, 25, 28,29,30,31,32], but their roles in the fetal testis are unknown. To define the protein expression and localisation of SOX7 and SOX17 during testis development, IF was used to examine SOX7 and SOX17 in testes collected from E12.5 and E15.5 embryos. SOX7 and SOX17 were both detected in CD31 positive endothelial cells within E12.5 and E15.5 testis, identifying them as definitive endothelial cell markers in the testis (Additional file 9: Fig. S4). Since SOX7 and SOX17 were co-expressed in CD31 positive endothelial cells and are known to act together in mediating vascular development in other tissues [32], the SOX7 and SOX17 antibodies were used in combination in subsequent experiments to increase sensitivity and ensure all SOX7/17 positive cells were detected. We next investigated E12.5, E13.5, E14.5, and E15.5 testes and found that SOX7/17 were detected in CD31 positive endothelial cells in the coelomic vessel and in endothelial cells between the testis cords (Additional file 10: Fig. S5).
RNA sequencing revealed that Sox7 and Sox17 transcription were 37.5- and 9.8-fold lower in testes treated with MEKi for 24h, suggesting Sox7 and Sox17 depend on MEK1/2 signalling for their expression in the testis. To confirm these data and determine whether SOX7 and SOX17 proteins were reduced in MEKi-treated samples, we also assessed SOX7/17 in E12.5 24h and 72h cultures using IF. Consistent with the E12.5-E15.5 IFs, CD31 positive endothelial cells located in the coelomic vessel and the mesonephros strongly expressed SOX7/17 in DMSO-treated samples cultured for 24h (Fig. 3A, Additional file 11: Fig. S6A-B). However, consistent with the RNA sequencing data, neither SOX7/17 nor CD31 positive endothelial cells were detected in testes treated with MEKi for 24h (Fig. 3A, Additional file 11: Fig. S6A). Notably, a small number of CD31 positive cells remained in the mesonephros of MEKi-treated samples and these remaining CD31 positive cells expressed SOX7/17 at substantially lower levels than in DMSO controls (Additional file 11: Fig. S6B).
Fig. 3
SOX7/17 expression depends on MEK1/2 signalling in the developing testis. Section or wholemount immunofluorescent imaging of E12.5 testes cultured for 24 or 72h with DMSO or 500nM of MEKi. A and C Section IF of cultured testes stained with DAPI (blue), SOX7/17 (red) and CD31 (endothelial cells and germ cells; cyan). B Three-dimensional stacked Z-series images of cultured testes showing Oct4-eGFP (germ cells; green), SOX7/17 (blue) and pERK1/2 (red). The DMSO and MEKi images should be viewed in conjunction with Movies S5-8 which reveal the 3D organisation of the cells in the tissue. Scale bar represents 100 μm. Arrows indicate SOX7/17 positive endothelial cells. Biological replicates; n = 3–4 per treatment
In addition, wholemount IFs of DMSO 24h controls revealed strong SOX7/17 staining co-localised with pERK1/2 in the celomic vessel, between the testis cords and in the mesonephros, but did not co-localise with Oct4-eGFP within the testis cords (Fig. 3B, Additional files 12–13: Movies S5-6). In 24h MEKi treated testes, pERK1/2 and SOX7/17 were not detected in the vast majority of cells, although very weak SOX7/17 staining was evident in a few remaining endothelial cells (Fig. 3B, Additional files 14–15: Movies S7-8). Together, these data suggest that SOX7 and SOX17 depend on MEK1/2 signalling in endothelial cells. Consistent with these findings, while CD31 and SOX7/17 double positive endothelial cells were detected in the coelomic vessel, between the testis cords, and in the mesonephros in 72h DMSO cultures, neither CD31, nor SOX7/17 were detected in the testis or mesonephros in 72h MEKi-treated samples (Fig. 3C, Additional file 11: Fig. S6C). Together, the RNA sequencing and IF data suggest that expression of SOX7 and SOX17 depend on MEK1/2 signalling in endothelial cells. However, it remains possible that SOX7 and SOX17 are lost simply due to endothelial cell death and further work is required to determine whether MEK1/2 signalling promotes Sox7 and Sox17 transcription in endothelial cells.
MEK1/2 signalling occurs downstream of VEGF in the developing testisVEGF signalling plays an important role in the establishment and patterning of vasculature in the testis [15]. Moreover, based on the DEGs found in our MEKi 24h and 72h RNA sequencing dataset, IPA identified VEGF/VEGFA signalling as potential upstream regulator(s) of MEK1/2 signalling in the testis. To test this, we collected E12.5 testes and cultured them for 24h in media containing 100, 500 or 2500nM of the VEGFR inhibitor, Cabozantinib (VEGFRi), and compared the outcomes with DMSO and MEKi treated testes. Using IF we assessed CD31 and pERK1/2 in endothelial cells at each dose. In the 24h DMSO control, CD31 positive endothelial cells were detected in the coelomic vessel and contained intense pERK1/2 staining (Fig. 4A, Additional file 16: Fig. S7A). In addition, consistent with our previous observations (Fig. 2, Additional file 3: Fig. S2), pERK1/2 was also detected in Sertoli cells at a lower intensity compared to endothelial cells (Fig. 4A, Additional file 16: Fig. S7A). At all VEGFRi doses, some CD31 positive endothelial cells were detected in the coelomic vessel region and the mesonephros. However, in contrast to the well organised vasculature found in DMSO controls, endothelial cells were not arranged in organised blood vessels in VEGFRi treated samples (Fig. 4A, Additional file 16: Fig. S7A), consistent with the established role for VEGRF in vascular patterning in the fetal testis. Although pERK1/2 was detected in CD31 positive endothelial cells following 100nM of VEGFRi, the intensity of pERK1/2 was substantially reduced compared to DMSO controls, indicating that 100nM VEGFRi partially inhibited VEGFR. Consistent with this, CD31 positive endothelial cells detected in gonads treated with 500 and 2500nM VEGFRi did not express pERK1/2 (Fig. 4A, Additional file 16: Fig. S7A). However, while pERK1/2 was not detected in endothelial cells, 100, 500 or 2500nM VEGFRi did not reduce pERK1/2 in Sertoli cells (Fig. 4A, Additional file 16: Fig. S7A). In addition, pERK1/2 was detected in surface epithelial cells of samples cultured with DMSO or with 100, 500 or 2500nM VEGFRi, although staining was less intense at 500 and 2500nM doses (Fig. 4A, Additional file 16: Fig. S7A). These data were confirmed in E12.5 gonads cultured for 24h with a second VEGFR inhibitor, Axitinib. While endothelial cells and vasculature were unaffected by DMSO, culture with 100, 500 and 2500nM Axitinib disrupted vascular formation and abrogated pERK1/2 expression in endothelial cells, outcomes similar to those observed using VEGFRi (Additional file 16: Fig. S7B).
Fig. 4
VEGFR is required for endothelial cell activation of pERK1/2 and vasculogenesis in the developing testis. Section and wholemount immunofluorescent imaging of E12.5 testes cultured for 24h with DMSO or 100, 500 or 2500 nM of VEGFRi. A IF in sections of cultured testes stained with DAPI (blue), pERK1/2 (red) and CD31 (endothelial cells and germ cells; cyan). B Three-dimensional stacked Z-series images of cultured testes showing Oct4-eGFP (germ cells; green), CD31 (endothelial cells and germ cells; blue) and pERK1/2 (red). The DMSO and VEGFRi images should be viewed in conjunction with Movies S9-12 which reveal the 3D organisation of the cells in the tissue. Notably, in the VEGFRi treatment this reveals a thin surface epithelium positive for pERK1/2, but negative for CD31 in the VEGFRi treated samples. Scale bar represents 100 μm. Arrows indicate endothelial cells and asterisks indicate pERK1/2 positive Sertoli cells. Biological replicates; n = 3–4 per treatment
E12.5 gonads treated for 24h with DMSO or 500nM of VEGFRi were also analysed using wholemount IF to assess Oct4-eGFP, CD31 and pERK1/2 (Fig. 4B). As previously observed (Fig. 2B, Additional files 4–5: Movies S1-2), pERK1/2 was detected in Oct4-eGFP negative cells within the testis cords in DMSO controls, consistent with pERK1/2 expression in Sertoli cells (Fig. 4B, Additional files 17–18: Movies S9-10). In addition, pERK1/2 was detected in CD31 positive endothelial cells throughout the testis, particularly within the coelomic epithelium, between the testis cords and in the mesonephros (Fig. 4B, Additional files 17–18: Movies S9-10). In VEGFRi treated testes, pERK1/2 was not detected in the majority of endothelial cells outside the testis cords but was detected in Oct4-eGFP negative cells within the testis cords, consistent with maintained pERK1/2 in Sertoli cells (Fig. 4B, Additional files 19–20: Movies S11-12). pERK1/2 was also detected in cells at the epithelial surface of the gonad and in the mesonephros, however, these cells were CD31 negative indicating that they were not endothelial cells (Fig. 4B, Additional files 19–20: Movies S11-12). Together, as VEGFRi blocked pERK1/2 in endothelial cells and disrupted organisation of these cells into blood vessels, these data suggest that VEGF signalling promotes MEK1/2 signalling in CD31 expressing endothelial cells and their organisation into testicular vasculature. In contrast, given that pERK1/2 was still detected in Sertoli cells of VEGFRi treated testes, activation of pERK1/2 in Sertoli cells appears to be independent of VEGF signalling.
SOX7/17 positive endothelial cells depend on VEGF signalling in the developing testisWe also assessed the expression of SOX7/SOX17 in VEGFRi testes to determine if endothelial cells expressing these key angiogenic transcription factors also depend on VEGF signalling. In DMSO controls treated for 24h, SOX7/17 was detected in CD31 expressing endothelial cells in the coelomic vessel (Fig. 5A, Additional file 21: S8A), as previously observed in DMSO controls for the MEKi experiments (Fig. 3A, Additional file 11: Fig S6A). Vasculature was substantially disrupted in testes treated with 100nm, 500nm or 2500nM VEGFRi for 24h, with fewer CD31 positive endothelial cells detected and those that were detected were not organised into recognisable blood vessels (Fig. 5A, Additional file 21: Fig. S8A). Moreover, SOX7/17 was undetected or almost undetectable in the remaining CD31 positive endothelial cells in 100nM VEGFRi treated samples and was undetected in 500 and 2500nM VEGFRi treated samples (Fig. 5A, Additional file 21: Fig. S8A). Moreover, CD31 positive endothelial cells were also detected in the mesonephros of VEGFRi treated gonads (Additional file 21: Fig. S8B), an outcome similar to that observed in MEKi treated gonads (Additional file 11: Fig. S6A-B). However, SOX7/17 was not detected or was substantially reduced in CD31 positive cells located in the mesonephros of VEGFRi treated gonads (Additional file 21: Fig. S8B). The observations in VEGFRi treated gonads were confirmed with 100-2500nM Axitinib, which eliminated SOX7/17 in CD31 positive endothelial cells and disrupted vasculature formation (Additional file 21: Fig. S8C). Together, as some CD31 cells remained in VEGFRi treated samples but SOX7/17 was lost, these data suggest that SOX7/17 expression depends on VEGF signalling in endothelial cells. Consistent with this and loss of VE-cadherin in gonads of samples treated with 500nM VEGFRi (Additional file 8: Fig S3), no CD31-pERK1/2 or CD31-SOX7/17 positive endothelial cells were detected in the gonad in samples treated for 72h with 500nM VEGFRi, despite robust detection of CD31 and pERK1/2 and CD31 and SOX7/17 in endothelial cells of control samples (Additional file 22: Fig. S9).
Fig. 5
SOX7/17 positive endothelial cells depend on VEGFR signalling in the developing testis. Section and wholemount immunofluorescent imaging of E12.5 testes cultured for 24h with DMSO or 100, 500 or 2500 nM of VEGFRi. A IF in sections of cultured testes stained with DAPI (blue), SOX7/17 (red) and CD31 (endothelial cells and germ cells; cyan). B Three-dimensional stacked Z-series images of cultured testes showing Oct4-eGFP (germ cells; green), SOX7/17 (blue) and pERK1/2 (red). The DMSO and VEGFRi images should be viewed in conjunction with Movies S13-16 which reveal the 3D organisation of the cells in the tissue. Scale bar represents 100 μm. Arrows indicate endothelial cells. Biological replicates; n = 3–4 per treatment
Wholemount IF analysis of Oct4-eGFP, SOX7/17 and pERK1/2 in E12.5 testes cultured with DMSO or VEGFRi for 24h produced similar results. In DMSO control testes, pERK1/2, SOX7/17 double positive cells were detected in the coelomic vessel, between the testis cords and in the mesonephros (Fig. 5B, Additional Files 23–24: Movies S13-14). In contrast, very few SOX7/17 positive cells were detected in the coelomic vessel, between the testis cords or within the mesonephros in VEGFRi treated testes, and the intensity of remaining SOX7/17 positive cells was substantially reduced compared to DMSO controls (Fig. 5B, Additional files 25–26: Movies S15-16). Moreover, the few SOX7/17 cells that were observed were negative for pERK1/2. Together, these data suggest that VEGF is required to mediate MEK1/2 signalling and ensure the presence of SOX7/17 expressing endothelial cells in the developing testis.
VEGF and MEK1/2 signalling are required for endothelial cell survivalGiven that there were no or very few CD31 positive endothelial cells detected in MEKi treated or VEGFRi treated testes respectively, compared to DMSO controls, we used IF to assess the cell death markers, cleaved Caspase3/9 (cCaspase3/9) together with 5-ethynyl-2’-deoxyuridine (EdU) incorporation to measure cell proliferation, and CD31 to identify endothelial cells. Flow cytometry was not
留言 (0)