
記住我






Allergens are compiled in few databases, being the World Health Organization/International Union of Immunologic Societies (WHO/IUIS) allergen nomenclature, the Allergome, the AllergenOnline, and the COMPARE among the most popular [17,18,19,20]. The WHO/IUIS allergen nomenclature database is probably the most widely used and frequently updated [21]. It provides information on allergen nomenclature, isoforms, variants and epitopes, protein biochemical name, species, protein molecular weight (MW), and routes of exposure, being organized according to major taxonomical groups and respective orders [17]. It also includes the links to databases on protein sequences (UniProt) and respective encoding genes (GenBank) [22, 23]. The other databases (COMPARE and AllergenOnline) also contain most of the referred information [24, 25]. The classification/inclusion of allergens into protein families can only be found in a different database, the AllFam [26]. This database catalogues the allergens into protein families, linking them to the WHO/IUIS Allergen Nomenclature and AllergenOnline databases, as well as correlating with data from the Pfam database [27]. However, its major gaps regard the absence of updated information since 2017 [26].
Currently, the WHO/IUIS allergen nomenclature lists 495 aeroallergens [17, 18], being divided in 103 families [26]. From those, 37% are distributed into 9 superfamilies of proteins, namely EF-hand superfamily (28 aeroallergens), profilin superfamily (28 aeroallergens), calycin superfamily (27 aeroallergens), prolamin superfamily (24 aeroallergens), subtilisin-like serine protease (21 aeroallergens), expansin superfamily (16 aeroallergens), Ole e 1-like proteins (16 aeroallergens), pathogenesis-related (PR-10) proteins, also known as Bet v 1 family (12 aeroallergens), and tropomyosins (11 aeroallergens), which are often subdivided in relevant groups containing important aeroallergens.
In this review, besides considering the number and importance of the allergens, the criteria of inclusion as inhalant protein families also considered their relevance as primary sensitizers of RA and their relationship with FA. PR-10 proteins and the nonspecific lipid-transfer proteins (nsLTP) (prolamin superfamily) are cases of important primary sensitizers of respiratory and food allergies in Europe.
EF-hand FamilyThe EF-hand family encompasses a variety of calcium-binding proteins disclosing a conserved motif with a helix-loop-helix secondary structure composed of a 12-residue calcium-binding loop flanked by 2 α-helices of 12 residues in length. EF-hand protein members tend to occur in pairs, meaning that most family members have two, four, or six calcium-binding motifs. This motif is crucial for eukaryotic cellular signaling, and the pairing enables communication, and positive cooperativity, reducing the Ca2+ signal needed to attain protein saturation [28]. The conformational effects of Ca2+-binding are diverse and depend on the function of the protein. EF-hand proteins display varying sensitivities to Ca2+, influenced by the intrinsic binding ability of the EF-hand and the degree of cooperativity in Ca2+-binding to paired EF-hands. Additionally, two factors can affect their ability to bind Ca2+, namely its selectivity towards Mg2+ (a divalent cation that is in a much higher cytoplasmic concentration) and its interaction with a protein target [28]. The biological functions of EF-hand family members encompass signaling and calcium buffering or transport processes.
This family is composed of different groups of proteins, some of them with allergenicity. The EF-hand superfamily ranks first position in terms of identified and listed aeroallergens, sharing this position with profilins [17]. Presently, 28 aeroallergens belonging to EF-hand family are divided into different subfamilies of proteins, namely polcalcins (17 allergens), troponin C (5 allergens), myosin light chain (MLC) (5 allergens), and sarcoplasmic calcium-binding proteins (SCBP) (1 allergen) [17]. Polcalcins are composed of aeroallergens from plant sources (e.g., Bet v 4 – European white birch), while troponin C, MLC, and SCBP groups contain mostly aeroallergens from animal sources (e.g., Der f 39 – American house dust mite, Bla g 8 – German cockroach, Bos d 3 – cow, respectively) (Table 1).
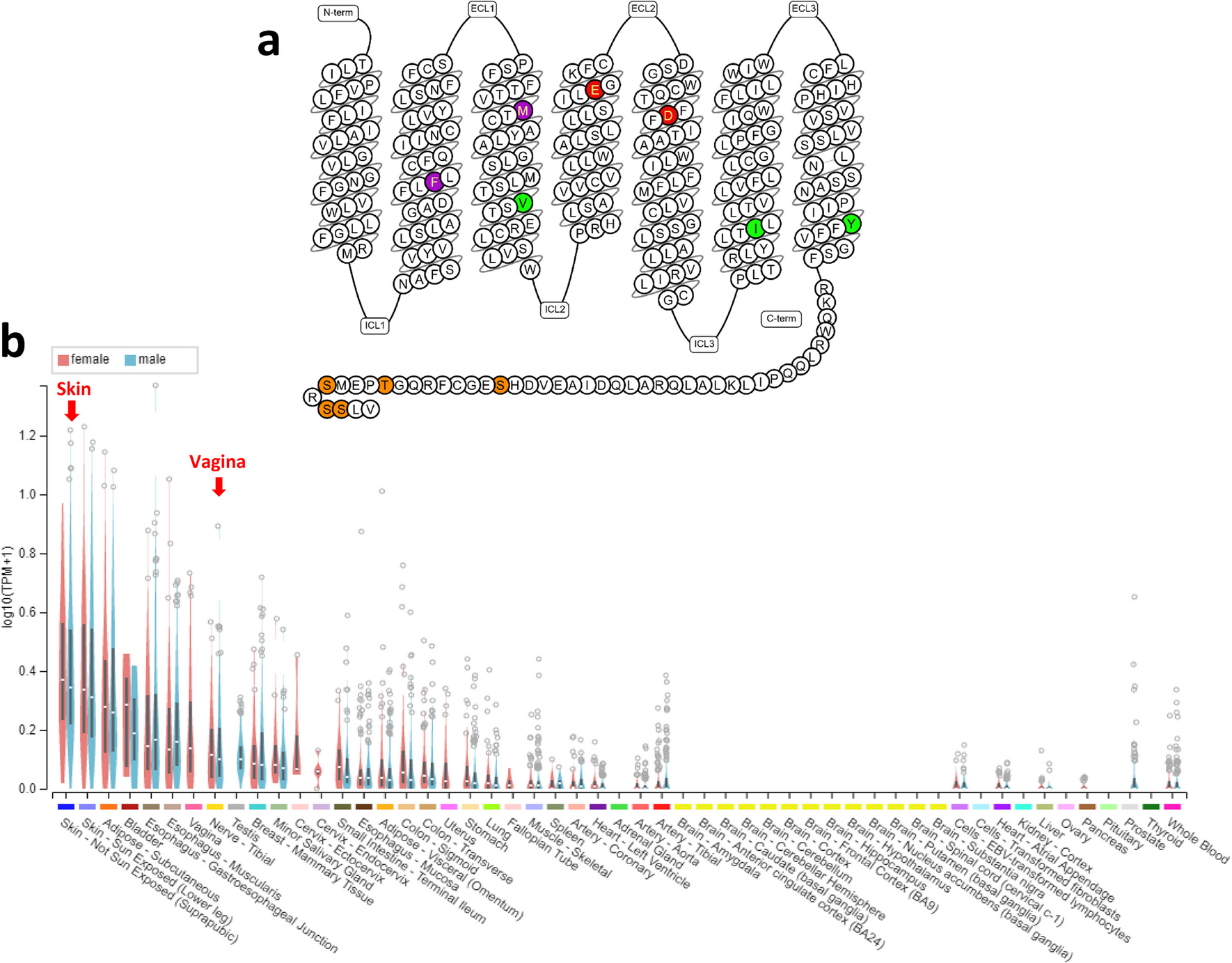
Table 1 Summary of physicochemical properties of selected aeroallergen families of proteinsPolcalcinsBased on the number of calcium-binding EF-hand motifs, polcalcins are categorized into three types, possessing different number of EF-hand motifs: two (e.g., Che a 3, white goosefoot), three (e.g., Bet v 3, European white birch), or four (e.g., Ole e 8, olive tree). The three-dimensional (3D) structure of polcalcins is characterized by α-helices forming a typical all-α protein fold (Fig. 1a, Table 1). The monomer, with a MW of 8–9 kDa, exhibits the characteristic polcalcin structural domain [29], but whose biological function remains unclear. However, given their specific localization in pollen and their calcium-binding ability, it is suggested that polcalcins may regulate intracellular calcium levels during pollen germination. Notably, the calcium-binding property of polcalcins influences their IgE-reactivity and thermostability [29]. Due to their high sequence identity and structural homology, these proteins are classified as panallergens [30]. Polcalcins are highly cross-reactive calcium-binding allergens specifically expressed in pollen tissues, meaning that sensitization to polcalcins is not associated with FA [29]. Less than 10% of allergic individuals are sensitized to polcalcins, so these proteins are classified as minor allergens [31]. The clinical expression of sensitization to polcalcins is still not clear, although symptoms are mostly described as mild. Patients sensitized to polcalcins often exhibit high cross-reactivity, as well as distinct IgE repertoires to different allergens, which highlights the need for monitoring these patients as they are susceptible of developing multiple pollen sensitizations [29]. So far, 17 polcalcins have been identified as aeroallergens in pollen (e.g. Ole e 3, olive) [17].
Fig. 1
Pymol representation of: a) Crystal structure of polcalcin allergen Che a 3 from Chenopodium album with calcium ions represented as green spheres (PDB ID: 2OPO); b) 3D structure of troponin C allergen Bla g 6 from Blattella germanica predicted by AlphaFold; c) 3D structure of myosin light chain allergen Bla g 8 from Blattella germanica predicted by AlphaFold; d) 3D structure of sarcoplasmic calcium binding allergen Aed a 5 from Aedes aegypdi predicted by AlphaFold; e) Crystal structure of profilin allergen Bet v 2 from Betula pendula (PDB ID: 5NZC); f) Crystal structure of lipocalin allergen Bla g 4 from Blattella germanica (PDB ID: 4N7C); g) Crystal structure of FABP allergen Der f 13 from Dermatophagoides farina (PDB ID: 2A0A); h) Crystal structure of nsLTP allergen Art v 3 from Artemisia vulgaris (PDB ID: 6FRR); i) 3D structure of α -amylase allergen Der p 4 from Dermatophagoides pteronyssinus predicted by AlphaFold; j) 3D structure of subtilisin-like protease allergen Cur l 4 from Cochliobolus lunatus predicted by AlphaFold; k) Crystal structure of β -expansin/Grass group-1 allergen Zea m 1 from Zea mays (PDB ID: 2HCZ); l) Crystal structure of Ole e 1-like allergen Lig v 1 from Ligustrum vulgare (PDB ID: 6YOA); m) Crystal structure of PR-10 allergen Bet v 1 from Betula pendula (PDB ID: 1BV1); n) 3D structure of tropomyosin allergen Der p 10 from Dermatophagoides pteronyssinus predicted by AlphaFold). All protein structures are represented in cartoon and colored by chain
Troponin CTroponin C is a calcium-binding protein that makes part of the troponin complex encompassing another 2 proteins, namely inhibitory (troponin I), and tropomyosin binding (troponin T). Troponin C is a muscle protein that connects the thin filaments and signals muscle contraction after binding Ca2+, presenting 2 structurally homologous lobes, mainly composed of α-helices and loops connected by a long uncovered 9-turn helical linker [32]. This protein plays a regulatory function, being common to animal sources. Sensitization to troponin C is often accompanied by cross-reactivity to its counterparts in foods, leading to an increased risk for individuals sensitized to this set of proteins [32]. So far, 5 troponin C have been classified as aeroallergens of insects/mites (e.g., Bla g 6, German cockroach) (Fig. 1b, Table 1), being classified as minor allergens [33].
Myosin Light ChainMLC comprises a multi-subunit complex of two heavy chains and four calcium-binding light chains. It serves as a fundamental contractile protein present in animal cells. MLC from invertebrates, including cockroaches and crustaceans, can bind IgE, being classified as aeroallergens and food allergens, respectively. So far, 5 MLC have been identified as aeroallergens in insects/mites (e.g., Bla g 8, German cockroach) (Fig. 1c, Table 1).
Sarcoplasmic Calcium-binding ProteinSCBP are classified as invertebrate muscle proteins exhibiting a calcium buffering function, being commonly considered as food allergens, especially in crustaceans. SCBP structure is greatly compact and globular with a hydrophobic core, contrarily to the dumbbell-shaped structure of troponin C [34]. So far, only one aeroallergen (Aed a 5) was identified in the yellow fever mosquito (Fig. 1d, Table 1). The clinical expression of this aeroallergen is still unknown, although it is considered a minor allergen [35].
Profilin SuperfamilyProfilins are a family of actin-binding proteins composed of 125–153 amino acids, with MW of 12–15 kDa and isoelectric points (PI) between 4.3–9.2 [36]. They have been characterized as monomers, although dimers and tetramers have also been reported for profilin structures [37]. These proteins are key regulators of actin polymerization in the cytoskeleton, participating in several cellular processes, including motility, metabolism, endocytosis, signal transduction, and gene transcription [38]. Profilins are ubiquitous in most eucaryotic cells, exhibiting high sequence identity (> 70%), and structural homology, even among distantly related family members, rendering their classification as panallergens [29] and presenting high levels of cross-reactivity among plant-allergenic sources (Table 1). However, the high structural homology between plant and mite profilins might not result in clinically relevant respiratory allergies, as there is no evidence of cross-reactivity between these 2 groups [39]. Bet v 2 (European white birch) pollen (Fig. 1e) was the first allergenic profilin to be identified in 1991 and since then, other allergenic profilins have been discovered in pollen, latex, and plant foods. Bet v 2 is constituted by a central 6-stranded antiparallel β-sheet and two helices, one at the amino-terminal side perpendicular to the sheet and another parallel at the C-terminal side.
Presently, profilins share the first place with EF-hand family in terms of the number of identified molecules, accounting for 28 profilins classified as aeroallergens [17]. Allergenic inhalant profilins are mainly found in plant pollens (e.g., Cro s 2, saffron) and mites (e.g., Tyr p 36, storage mite), being important triggers of RA (e.g., allergic asthma) and relevant primary sensitizers of FA. Profilins have been recognized as clinically relevant aeroallergens, with sensitized/allergic subjects clinically reacting to multiple allergen sources [40]. Typically, profilins are associated with mild respiratory symptoms, but recent data seem to highlight their importance as causative agents for severe/systemic allergic reactions like anaphylaxis, especially in FA [40]. In Europe, 20–30% of individuals allergic to pollen are IgE-reactive to profilins [41, 42], leading to a minor allergen classification. However, there is no correlation between the level of IgE-reactivity of individual patients to specific profilins and their sensitizing source, signifying that sensitization could be linked to any aeroallergenic profilin. This fact highlights the usefulness of profilins as a common marker for polysensitization for diagnosis and therapeutic approaches [43].
Calycin Protein SuperfamilyThe calycin superfamily is widely dispersed throughout all kingdoms and gathers a miscellaneous group of proteins known for their ability to bind small hydrophobic molecules. These proteins are involved in a variety of biological processes, including transport and storage of lipids, regulation of gene expression, and immune responses. The calycin core fold encompasses an eight-stranded calyx-shaped antiparallel β-barrel, which forms a hydrophobic binding cavity [44]. This calycin superfamily is composed of different families of allergenic proteins, namely lipocalins and fatty acid-binding proteins (FABP), ranking the second position in terms of the total number of identified aeroallergens (27 proteins) (Table 1) [44].
LipocalinsLipocalins are a diverse group of proteins with multiple biological roles that include (i) the transport of small hydrophobic molecules and (ii) the regulation of various metabolic, development, and immunological processes [45]. Lipocalins present 150–250 residues and a MW of 17–25 kDa, being extracellularly located. Although the sequence identity among lipocalins is generally low (20–30%), it can be higher than 50% in some cases, and with well-preserved 3D structures, sharing an eight-stranded antiparallel β-barrel motif that forms a ligand-binding pocket, and an α-helix (Table 1). Allergenic lipocalins sensitize efficiently, with an average sensitization rate > 50%. Presently, 21 lipocalins are classified as aeroallergens, including allergens like Bla g 4 (cockroaches) (Fig. 1f), Mus m 1 (mouse urine), and Can f 1 and Can f 2 (dog), most of them behaving as major allergens. Lipocalins are predominantly abundant in mucosa and skin epithelia, being extensively observed in body fluids and secretions, which makes them prevalent in indoor aeroallergens [17].
Fatty Acid-Binding ProteinsCytosolic (cytoplasmic) fatty acid-binding proteins (FABP) are a family of proteins primarily located in the cytosol, playing a crucial role in lipid metabolism by acting as intracellular transporters of hydrophobic metabolic intermediates and as carriers of lipids between membranes. FABP exhibit significant sequence and structural similarity, being small soluble proteins with 12–18 kDa and 110–160 residues. They are typically monomers but with tendency to form self-associated structures [46]. These proteins have a mixed α-β structure, with a classic β-barrel made up of 10 antiparallel β-strands and covered with a pair of α-helices at one end, creating an internal cavity that serves as the ligand-binding site [47, 48]. Ligand specificity and affinity are influenced by the side chains of amino acids that extend into the cavity, particularly polar residues that engage in hydrogen bonding and electrostatic interactions with the polar head groups of the ligands. For instance, arginine binds fatty acids, while glutamine binds retinoids. The antiparallel β-barrel fold is also common to lipocalins, which similarly bind small hydrophobic molecules. However, sequence similarity between FABP and lipocalins is limited to a short N-terminal motif. Presently, 6 FABP are identified as aeroallergens, all belonging to mites (e.g., Der f 13, American house dust mite (HDM)) (Fig. 1g, Table 1) [17].
Prolamin SuperfamilyProteins of the prolamin superfamily are composed of a high content of glutamine and proline residues, which is the typical trace of prolamins. Members of this superfamily show little sequence homology, although preserving a common cysteine skeleton and α-helical structures [49]. They have a preserved pattern of eight cysteine residues, being responsible for their 3D structure stabilization, presenting a right-handed super-helix formed by four α-helices. Prolamin superfamily includes several protein families, but their identified aeroallergens are mainly included in the nsLTP and α-amylase families. Presently, this superfamily occupies the third place in terms of identified aeroallergens, totalizing 24 molecules [17].
nsLTPThe nsLTP constitute a large family of proteins that are abundant in all plants. Most nsLTP are extracellular proteins associated with cell walls, possessing lipid-binding specificity in their 3D structure. nsLTP are also known as the PR-14 family, whose members can intervene in the modulation of lipid composition, lipid transfer, vesicular trafficking, and signal transduction. nsLTP are small proteins with 9.5–10.5 kDa, presenting compact 3D structures [50], consisting of four α‐helices, which are stabilized by four highly conserved disulphide bridges and connected by flexible loops [51], forming an internal, tunnel‐like, hydrophobic cavity, responsible for the transport of various lipids. Allergic reactions triggered by inhalant nsLTP can range from mild to potentially life-threatening, such as anaphylaxis [52]. nsLTP are classified as panallergens due to their high homology and wide presence among plant species. Presently, 16 nsLTP have been identified as aeroallergens (e.g. Amb a 6, ragweed) (Fig. 1h) in pollens and leaves (Table 1) [17]. For several years, nsLTP-sensitized allergies were more prevalent in the Mediterranean region, but currently, this paradigm has changed with nsLTP progressively functioning as primary sensitizers in other geographical areas [51, 53, 54]. The nsLTP are related to important clinical conditions, such as LTP syndrome and pollen-food allergy syndrome (PFAS) [55].
α-Amylasesα-Amylases are metalloenzymes found in distinct glycoside hydrolase families, being ubiquitously distributed. They are present in different sources, namely microorganisms, plants and animals, displaying biological functions that regard the catalysis of α-1,4-glucosidic bonds in starch and related α-glucans with retention of α-anomeric configuration in the products [56]. The amylolytic digestion produces glucose, maltose, and maltodextrins. In addition, many α-amylases are also able to catalyze transglycosylation. Structurally, α-amylases encompass a single polypeptide chain folded into three domains (A, B and C). These enzymes present a characteristic (β/α)8-barrel with conserved catalytic core (domain A), a protrusion between the third strand and the third helix of (β/α)8-barrel having irregular β-like structure (domain B) and a C-terminal end of the amino acid sequence with key motif (domain C) (Table 1) [57].
Presently, 8 α-amylases have been identified as aeroallergens in mites (e.g., Der p 4, European HDM) (Fig. 1i), insects (e.g., Bla g 11, German cockroach), and fungi (e.g., Asp o 21, rice mold). α-Amylases have been described to cause occupational asthma due to the inhalation of flours and powders commonly present at bakeries, as well as in pharmaceutical and laboratory facilities [58, 59]. More recently, α-amylase from yellow mealworm has also been reported to be the culprit aeroallergen-inducing occupational allergy in workers contacting with mealworms in different circumstances, namely in pet stores, live fish bait or infested stored grains, as well as in mealworm farming for animal feed and human consumption [60].
Subtilisin-like Serine ProteaseSubtilisin-like proteases (subtilases) are a family of serine proteases exhibiting a characteristic catalytic triad consisting of aspartate, histidine, and serine [61]. These enzymes have a unique structural fold, comprising a highly twisted seven-stranded β-sheet flanked by two layers of α-helices. Subtilases are typically monomers although homodimerization can also occur [62]. Their MW varies from 34–58 kDa [17], depending on the location of the enzymes (vacuolar versus extracellular) (Table 1). Their physiological role depends on the allergen source, participating in the production of nutrients (through protein cleavage), protein turnover, fungi/plant defense system, and immune evasion [63]. Allergenic subtilases can alter the epithelial integrity through degradation of tight or adherens junctions, thus facilitating their penetration into submucosal tissues. Their contact with immune cells, facilitated by the epithelial barrier degradation, allows the cleavage of surface receptors on target cells, which optimizes the Th2-biased inflammatory response. Presently, it is the fourth family gathering more aeroallergens, totalizing 21 subtilases belonging to different sources, namely fungi (e.g., Cur l 4, mold) (Fig. 1j) and mites (e.g., Per a 10, American cockroach). Allergic symptoms associated with subtilisin sensitization may include breathlessness, sweating, and wheezing at exposures, contributing to occupational asthma [64].
Expansin SuperfamilyThe expansin superfamily encompasses a set of secreted proteins (25–27 kDa) expressed in plants and divided into four families: α-expansin, β-expansin, expansin-like A, and expansin-like B. Current identified aeroallergenic β-expansins are included in the commonly known Grass group-1 allergens. Their biological role is related to cell wall loosening processes, thereby aiding in the pollination process [
留言 (0)