
記住我

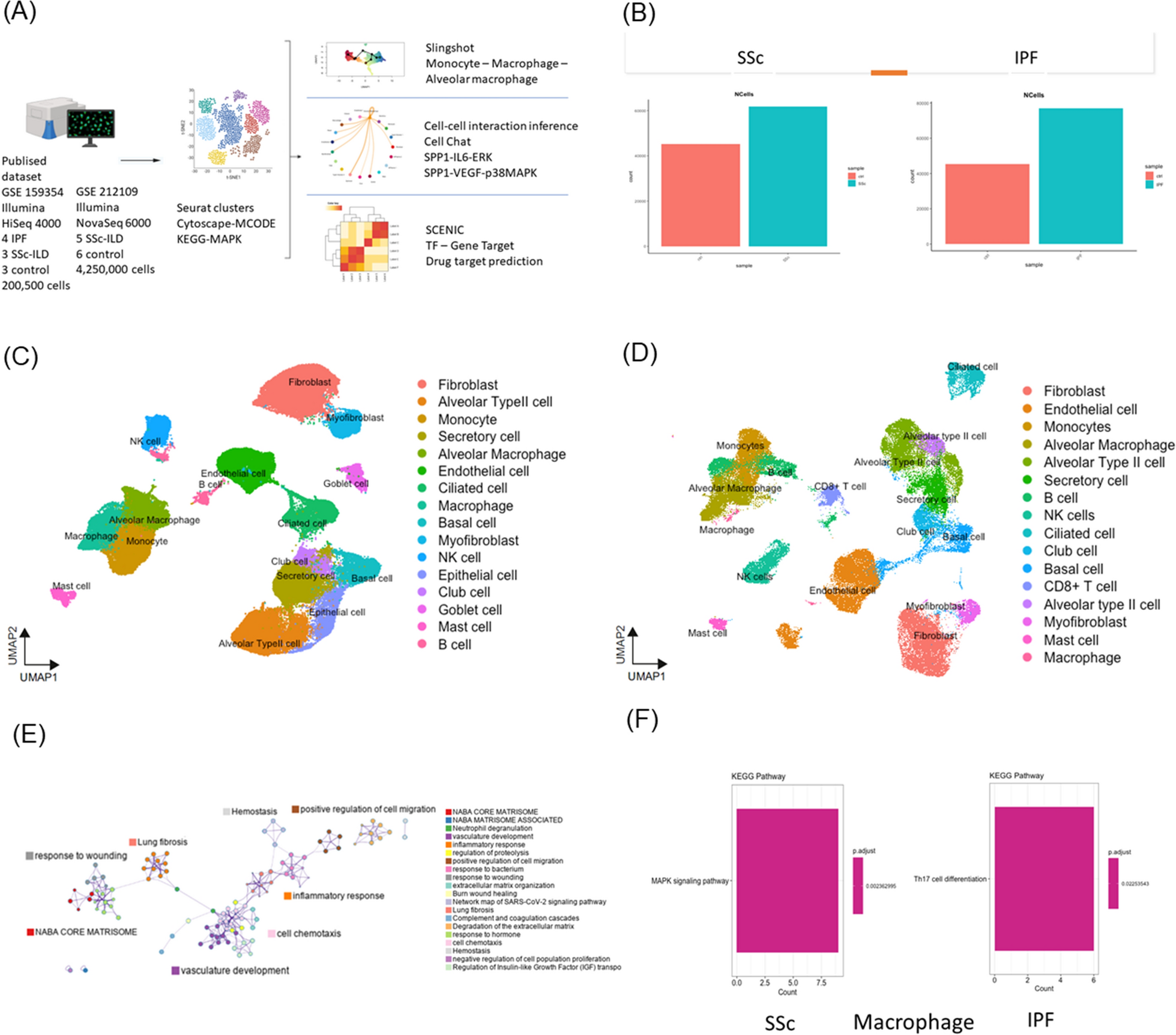


We analyzed lung samples from four patients with IPF, three patients with SSc-ILD patients, and three HCs from GSE159354 using scRNA-seq analysis (Fig. 1A). The filtered genes were integrated with batch corrections using the Seurat software. We conducted a bioinformatics analysis of samples in two groups of patients with SSc-ILD or IPF with HCs. In total, approximately 200,500 cells were analyzed (Fig. 1B). A total of 19 and 23 clusters were identified in the SSc-ILD with HCs (Fig. 1C and Fig S1B, C) and IPF with HCs groups (Fig. 1D), respectively. Based on the marker genes that were used to annotate and identify the clusters, the cell clusters were similar in both groups (Fig. 1C and D).
Fig. 1
Single-cell RNA-sequencing analysis of lung samples from sysetmic sclerosis (SSc), idiopathic pulmonary fibrosis (IPF) and healthy controls (HCs). A Overview of study design and analysis. B–F Overview of the scRNAseq landscape of GSE159354. Markers were used to identify the clusters and differences among the SSc-ILD, IPF, and HC groups. B The analyzed cell counts of the SSc-ILD, IPF, and HC groups. UMAP of samples from the (C) SSc-ILD and HC group and (D) IPF and HC group. E Enriched ontology clusters of all samples. F Bar plots showing the KEGG pathways of macrophages in SSc-ILD and IPF. Illustrations in A were created using BioRender (http://biorender.com)
Lung macrophages execute MAPK signaling pathway specific for SSc-ILDIn the enriched ontology clusters, we observed two cluster groups associated with lung fibrosis: the inflammatory process and complement cascade, and the wound healing process (Fig. 1E). Because of the similar clustering patterns and GO exhibited by these two sets of samples (SSc-ILD and IPF) (Fig S1D, E), we conducted an MCODE component analysis (Fig S1F). We identified the mitogen-activated kinase phosphatase (MAPK) signaling pathway in SSc-ILD, but not in IPF (Fig S1F). Therefore, we aimed to determine which group of cells was executing MAPK. KEGG analysis on fibroblasts, epithelial cells, monocytes, alveolar macrophages, and macrophages revealed that macrophages executed MAPK (Fig S1G–J, Fig. 1F); in contrast, the same group of macrophages in IPF underwent Th17 cell differentiation (Fig. 1F).
Monocyte-derived lung macrophagesIn the three Seurat clusters representing monocytes/macrophages of clusters 2, 4, and 7, the differentially expressed genes (DEGs) included FCN1, S100A8, and IL1B in cluster 2; APOC1, MARCO, FABP4, and SPP1 in cluster 4; and HLA-DPB1, GPR183, and CCL3 in cluster 7 (Fig. 2A). According to the CellMarker 2.0 database annotation [16], clusters 2, 4, and 7 corresponded to monocytes, alveolar macrophages, and macrophages, respectively (Fig. 2B). To determine the origin of the macrophages within these clusters, we performed slingshot [20] to distinguish the similarities between these three cell types. Starting with monocytes, we found that, to reach alveolar macrophages, one must first pass through the macrophages. There were distinct differentiation pathways from monocytes to alveolar macrophages and macrophages (Fig S1K). Thus, macrophages are more closely related to monocytes, whereas alveolar macrophages likely arrive in the lung tissue earlier and undergo more differentiation, resulting in notable differences from monocytes. Consequently, we concluded that macrophages were more likely to differentiate from monocytes (Fig. 2C).
Fig. 2
Single-cell RNA-sequencing analysis of monocyte/macrophage from systemic sclerosis (SSc). A–C Overview of the scRNAseq landscape of GSE159354. A Differential expression of key gene heatmap across cluster 4-alveolar macrophage, cluster 2-monocyte, and cluster 7-macrophage of SSc-ILD. B Violin plots showing the differential expression of key genes across cluster 2-monocyte, cluster 4-alveolar macrophage, and cluster 7-macrophage of SSc-ILD. C Suggested trajectory from monocytes, macrophage, and alveolar macrophages of SSc-ILD on the 2D map. D, E Overview of the scRNAseq landscape of GSE212109. Markers were used to identify the clusters and differences between SSc-ILD and HC. D UMAP of samples from the SSc-ILD and HC group. E Violin plots showing the differential expression of key genes across clusters 1, 3, 12, and 26
The alveolar macrophage subpopulation in SSc-ILDUsing the published dataset GSE 212109, we analyzed lung samples from five patients with SSc-ILD and six HCs (Fig. 1A). In the scRNA-seq analysis, 27 clusters were identified (Fig. 2D). Based on the marker genes used for annotating clusters and assigning identifications, clusters 2 and 7 corresponded to monocytes, cluster 8 to macrophages; and clusters 1, 3, 12, and 26 to alveolar macrophages (Fig S1L). In the alveolar macrophage subset, cluster 1 consisted of cells expressing markers for both FABP4 and MARCO, clusters 3 and 26 represented cells expressing SPP1, and cluster 12 comprised cells expressing both FABP4 and SPP1 (Fig. 2E). This observation aligns with the published dataset GSE159354, as we identified two groups of cluster 4-alveolar macrophage, one with the key genes FABP4 and MARCO, and the other with MARCO and SPP1 (Fig. 2A).
Cell–cell interaction inference (ligand–receptor interaction)Given the differences in gene expression, our focus shifted to understanding how intercellular interactions may contribute to SSc-ILD and potentially lead to earlier disease onset compared to IPF [8]. Through functional differences between cell cluster interactions for candidate signaling pathways, we initially observed that type II alveolar cells, epithelial cells, secretory cells, and goblet cells tended to produce UGPR1, SAA, and complement signaling pathways upon injury (Fig. 3A, B, Fig S2A, B). UGPR1 activates alveolar macrophages bearing MARCO (Fig. 3C) [26], whereas the complement signaling pathway influences alveolar macrophages to drive additional immune and repair responses (Fig. 3D). In contrast, SAA acts on monocytes and triggers subsequent inflammatory reactions (Fig. 3E) [27].
Fig. 3
Functional differences between cell cluster interactions for candidate signaling pathways. A River plots of outgoing and incoming signal between cell clusters in SSc. B Heatmap highlighting the differential cell interaction strengths of outgoing and incoming signals. C–J Plots showing cell–cell interaction and strength for specific pathways, including (C) UGPR1, (D) COMPLEMENT, (E) SAA, (F) SPP1, (G) BAFF, (H) VEGF, (I) VISFATIN, and (J) IL6. K Heatmap illustrating cell–cell interaction and strength of ANNEXIN signaling pathway. L Graphical abstract of the lung single-cell interaction map of SSc-ILD. Illustrations were created using BioRender (http://biorender.com)
Subsequently, alveolar macrophages produced SPP1 (Fig. 3F, Fig S2C) and BAFF (Fig. 3G). SPP1 not only acts on fibroblasts and epithelial cells for immediate repair (Fig. 3F), but also targets monocytes, further initiating an alternative repair pathway through the VEGF (Fig. 3H) and VISFATIN signaling pathway (Fig. 3I). Within monocytes, a distinct subset of macrophages executes this signaling pathway by specifically interacting with endothelial cells (Fig S2D, E). BAFF interacts with B cells to produce autoantibodies that are believed to be related to the autoimmune response [28].
We also observed that IL6, generally considered an inflammatory cytokine, was not produced by typical immune or inflammatory cells. Instead, it is generated by fibroblasts, myofibroblasts, and endothelial cells, and it acts on epithelial cells to facilitate repair processes (Fig. 3J). We also found that many cells, especially mast cells, produce ANNEXIN, which primarily acts on monocytes and alveolar macrophages, without affecting macrophages (Fig. 3K). ANNEXIN inhibits inflammatory responses via formyl peptide receptors (FPRs) [29]. However, macrophages lacked the annexin-FPR axis signaling pathway (Fig S2F). When macrophages are not inhibited by ANNEXIN, this may be the reason for the continued action of VEGF and VISFATIN.
We found that mast cells play a pivotal role in the feedback inhibition of monocytes. In previous studies, monocytes were found to possess two annexin receptors, FPR1 and FPR2. FPR1 primarily exhibits anti-inflammatory effects, while FPR2 aids the differentiation of monocytes into macrophages for efferocytosis [30]. However, in SSc-ILD, once monocytes differentiate into macrophages, both FPR1 and FPR2 disappeared (Fig S2F), consequently eliminating feedback inhibition from mast cells on macrophages. In contrast, monocytes in IPF possess only FPR1, and after differentiation into macrophages, the remaining FPR1 appears. Therefore, in IPF, feedback inhibition by mast cells persisted among monocytes, alveolar macrophages, and macrophages (Fig S2G).
The biological pathway and pathogenesis of SSc-ILDThe biological pathway of SSc-ILD is outlined in Fig. 3L. When frontline cells in the lung tissue such as type II alveolar, epithelial, secretory, and goblet cells are exposed to external or internal damage, they initially send UGPR1 signals to alveolar macrophages. Consequently, alveolar macrophages activate fibroblast and other cells, such as myofibroblasts and endothelial cells, through SPP1. Endothelial cells can act as a source of myofibroblasts through the endothelial-to-myofibroblast transition (EndMT) and concomitant microvascular rarefaction [31]. Subsequently, through the IL6, ERK, and pI3k-Akt pathways [32], they drive cells such as epithelial cells and fibroblasts to continue repairing the injured tissue. The repair process contributes to lung fibrosis.
Additionally, SPP1 produced by alveolar macrophages drives monocytes, and within the recruited monocytes, some differentiate into specific macrophages activating the VEGF and VISFATIN signaling pathways due to the hypoxic characteristics of SSc, further driving the downstream p38-MAPK pathway [28, 33]. This process contributes to the development of lung fibrosis.
While there are mechanisms in place to inhibit alveolar macrophages and monocytes, especially by mast cells through ANNEXIN to suppress inflammation [34], this feedback mechanism does not act on macrophages. As a result, inflammation caused by oxidative stress cannot be inhibited, leading to a vicious cycle and an early accelerated onset of inflammation-induced lung fibrosis. Alveolar macrophages also drive B cells to produce autoantibodies through BAFF, which may continuously affect autoimmunity [35].
p38-MAPK and JUN of the MAPK signaling pathway in lung macrophageWe observed that this specific subset of macrophages activating the MAPK signaling pathway led to an earlier onset of SSc-ILD. SCENIC was used to predict TFs and putative target genes (Fig S3). We found two regulon groups: the first group included BCLAF1, IRF1, and NFE2L2, and the second group included JUN, FOS, and FOSB (Fig. 4A). From these two regulon groups, we identified that the target genes of each TF belonging to the MAPK signaling pathway (Fig. 4B). Within the TF and target gene networks, BCLAF1 regulates IRF1, NFE2L2, and JUN. Additionally, JUN is regulated by FOS and FOSB, which influence downstream genes (Fig. 4C). These two groups of genes were related to the p38-MAPK and JUN pathways (Table S1).
Fig. 4
Regulons (transcription factor and downstream target genes) identified using SCENIC. A Heatmap of upregulated regulons of macrophages in samples of SSc-ILD. The AUC and binary matrix. B The upregulated regulons of macrophages overlapping with the MAPK pathway. C Networks of transcription factors and downstream target genes
Drug targeting prediction of transcription factorsMacrophage TFs are important targets of the MAPK signaling pathway. We mapped drugs targeting these TFs using the DrugBank database (https://go.drugbank.com/) (Table S2) [23]. Among the potential drugs, we selected those with downregulated effects on TFs and excluded those with controversial outcomes or those causing upregulated effects on other TFs. Additionally, we narrowed down the selection to those with an adjusted p-value < 0.05 (Table 1). Finally, we found that metformin downregulated NFE2L2. Metformin is a biguanide drug that is used to treat type 2 diabetes. It also has advantages in other diseases, including cancers and liver and renal diseases [36]. Previous reports suggest that metformin can inhibit the phosphorylation of Raf and ERK in a dose-dependent manner, thereby further suppressing NFE2L2 expression [37, 38].
Table 1 The drug scores of down regulated effect drugs to specific transcription factors
留言 (0)