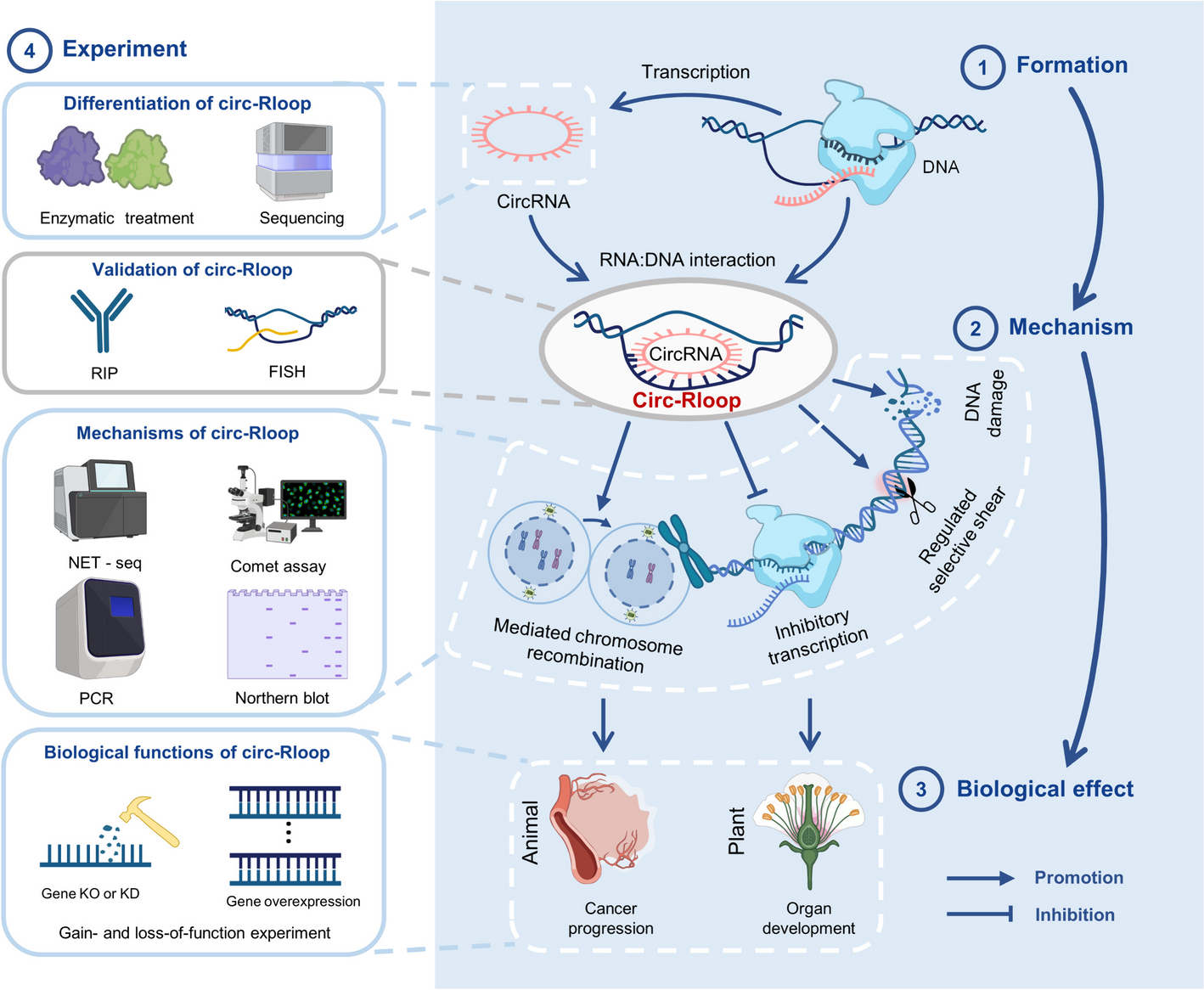
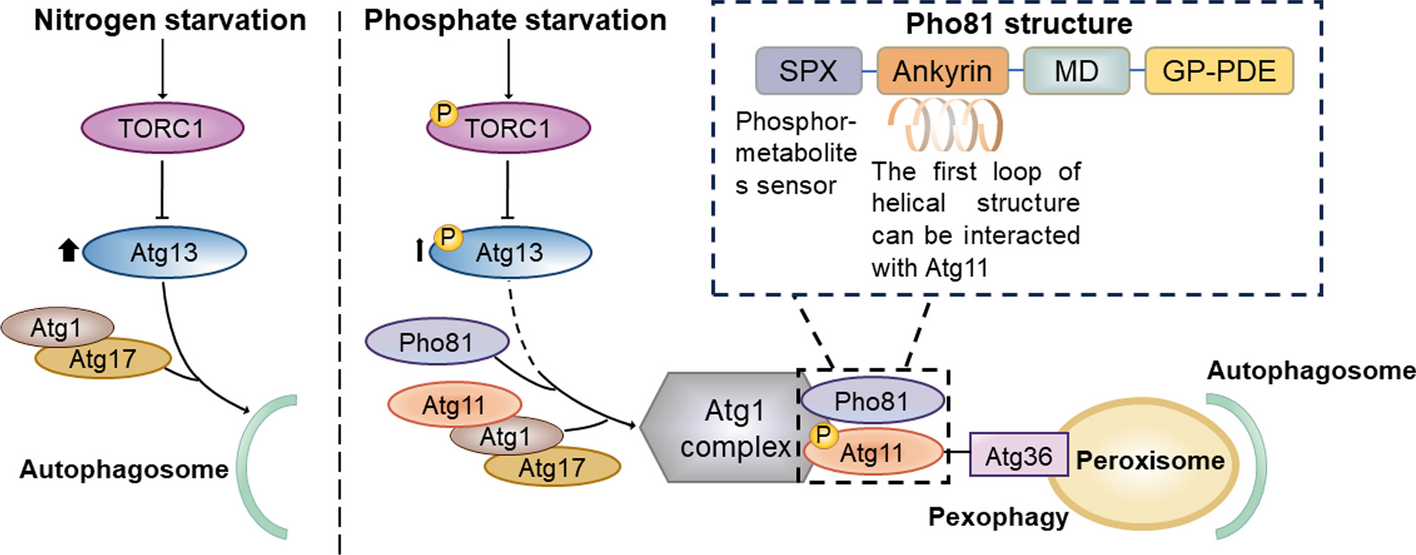
Identification and characterization of circR-loop
As a universal and extensively studied regulatory mechanism, the structure and function of R-loops have been thoroughly explored. However, the discovery of circR-loop, formed through interactions between circRNA and DNA, is a relatively recent development, with only a limited number of dedicated studies to date [13, 16]. This scarcity of research may be attributed to the challenges in identifying and distinguishing circR-loop. While circRNA was observed as early as 1979, its unique structure made it challenging to identify, often leading to confusion with linear RNA or outright dismissal as an aberrant sequence. It wasn't until 2013, with advancements in sequencing technology, that circRNA gained recognition and widespread study [26, 27]. Currently, research on circR-loop faces similar obstacles, as distinguishing circR-loop from other R-loops primarily relies on RNase H and RNase R processing or the speculative overlap of genomic sites. Novel methods for isolating, purifying, amplifying, and stabilizing circR-loop have yet to emerge, and standardized identification and classification criteria remain underdeveloped.
It is heartening to witness the development of breakthrough characterization technologies, which often offer superior genome coverage and enhanced detection accuracy compared to traditional methods. A notable example is the recent development of Cross-linked Poly(A) Pulldown RNase R sequencing (CLiPPR-seq) technology by Amaresh C. Panda and colleagues. This innovative approach employs 4′-aminomethyltrioxsalen hydrochloride (AMT)-mediated RNA-RNA double-stranded cross-linking treatment for enriching circular RNA and subsequent deep sequencing. By applying CLiPPR-seq, hundreds of circRNAs interacting with mRNA were identified across three different intact cell types, including βTC6, C2C12, and HeLa [28]. The continued refinement of this pioneering technique holds promise for uncovering unexplored circR-loops that potentially regulate gene expression.
The progression of sequencing technology is pivotal for high-throughput screening and identification of circR-loop, and future research must establish classification standards and innovative detection techniques to facilitate comprehensive mapping, laying the groundwork for subsequent investigations. Additionally, a more practical avenue for improvement involves integrating existing technologies and designing approaches based on specific sites or structures involved in the biological processes of R-loop formation and resolution. This approach would target genomic single-stranded DNA (ssDNA) exposure using techniques such as KAS-seq, CUT&TAG, and others, which would be combined and enhanced with circRNA characterization technology. This especially includes long-read sequencing detection based on third-generation sequencing technology, allowing for precise localization of circR-loop structures.
Establishing a standardized nomenclature for circR-loop
Standardization of the naming of newly discovered structures is crucial for promoting academic exchange and cooperation. However, due to limited research, there is currently a lack of unified nomenclature and classification standards for the R-loops formed by this type of circRNA, further complicating its definition and classification. Therefore, we propose the use of "circR-loop" as a unified name for this special structure. The prefix "circ" signifies that the RNA molecule involved is circRNA with a covalently closed circular structure, imparting unique stability and functional properties compared to linear RNA. The "R-loop" section emphasizes the distinctive hybrid structure formed between the RNA part and DNA, a phenomenon well-studied and confirmed previously [29, 30].
The origin, production and degradation mechanism of circR-loop
Meanwhile, critical biological properties such as the origins and mechanisms of action of circR-loop remain largely uncharted territory. While the formation mechanism and influencing factors of circR-loop are assumed to parallel those of traditional R-loops, specific mechanisms have yet to be uncovered and elucidated. Notably, while circR-loop structure has been explored in the mitochondrial genome, its structure within the nucleus remains unexplored. Given its status as an RNA–DNA interaction, the nuclear structure of circR-loop may possess broader relevance. Understanding the origin and structure of circR-loop is crucial, as it can shed light on the specificity of their regulatory functions both within and between species, warranting in-depth exploration as a priority.
As a type of ncRNA, circR-loops are theoretically formed by the hybridization between circRNA and distant DNA strands through trans interaction [1]. However, the specific formation process and mechanism of circR-loops are still not fully characterized. Future research can focus on the unique spatial structure of circRNA and R-loops, exploring the characteristic local chromatin 3D conformation and sequences (such as Alu and its derivatives) before circR-loop formation. This investigation will shed light on the novel mechanism underlying the formation of this specialized R-loop structure [31,32,33].
Moreover, the stabilization and degradation processes following circR-loop formation are also of great interest. Due to its covalently closed-loop structure, circRNA is typically more stable than linear RNA, resisting degradation by RNases and clearance by the body's natural immunity [34]. Various factors, including endonuclease activity, structural changes, and chemical substances, participate in circRNA degradation [35]. Conversely, the stability of R-loops is known to be influenced by factors such as transcription, GC content, and the presence of G-quadruplex structures [36,37,38]. Additionally, RNA binding and chromatin modification factors, along with helicases/translocases, topoisomerases, and replication/repair factors, collaborate to either form or eliminate R-loops [39, 40]. Given that nuclear accumulation of circRNAs can promote the formation of deleterious R-loops [17], further exploration of the degradation mechanism of circR-loops and associated components using advanced characterization technologies, such as microfluidics and sequencing, is essential. Understanding these mechanisms is crucial for developing related technologies to maintain cellular homeostasis and facilitate the biological functions of circR-loops [35].
CircR-loop’s unique and complex regulatory functions
Currently, the regulatory mechanism of circR-loop remains inadequately studied. Previous research has shown that regulatory R-loops play a crucial role in maintaining genome stability by modulating transcription activity, replication, recombination, centromere function, and DNA editing [29]. Additionally, circRNAs have been identified as regulators of gene expression through mechanisms such as miRNA or protein sponge adsorption and interaction with synthetic peptides [34]. Importantly, circRNAs can independently regulate genome stability or collaborate with other proteins in this process [34]. Consequently, it is anticipated that future research will focus on elucidating the role of circR-loop in regulating genome stability. Furthermore, given the widespread distribution of circRNAs and R-loops, it is likely that circR-loops are involved in regulating downstream gene expression through diverse mechanisms. Additionally, owing to the exceptional stability of circRNA, its regulatory mechanisms and capabilities likely diverge from those of other R-loops, necessitating further profound inquiry. While the intrinsic regulatory function of R-loops operates at the epigenetic level, the regulatory mechanisms of circR-loop, although conceptually akin, may entail unique attributes. Future research should initially develop methods tailored to the structure and characteristics of circR-loop based on existing experiments, thereby distinguishing the specific regulatory functions of circR-loop. Furthermore, when investigating circRNA function, circR-loop should be considered as a potential regulatory mechanism, extending beyond the study of ceRNA mechanisms.
The potential role of circR-loop in human disease
The potential impact of R-loops and circR-loops in human diseases cannot be underestimated. Dysregulated dynamics of R-loops formed by multiple linear RNAs have been implicated in DNA damage and genome instability, closely associated with diseases like cancer and autoimmune disorders [37]. In cancer, R-loop accumulation, influenced by various factors, drives DNA damage and replication stress [41, 42]. In autoimmune conditions such as Aicardi-Goutières syndrome (AGS), R-loops may contribute to disease pathogenesis by altering gene expression or reactivating retromers [43]. circRNAs have been implicated in the regulation of diverse diseases, including cancer, cardiovascular, neurological, and infectious diseases [8]. Although the regulatory role of circR-loops has been predominantly studied in cancer, with less exploration in other diseases, a recent study demonstrated that circLrch3 forms a circR-loop by binding to specific DNA sequences within the promoter of its host gene Lrch3. This promotes chromatin activation and DNA demethylation, enhancing Lrch3 transcriptional activity and leading to apoptosis of pulmonary artery smooth muscle cells [44]. Given the broad regulatory potential of circR-loop components in human diseases, further exploration across various disease types and sample cohorts is warranted to expand our understanding of the functional landscape of circRNA.
The potential of circR-loop in clinical translational research
The distinctive expression profile of circRNA presents significant potential as biomarkers, therapeutic agents, and drug targets. As a subtype of circRNA, circR-loop possesses a distinct spatial and sequence structure, offering broad clinical applications. However, clinical translation research on circR-loop remains nascent. Future exploration should focus on elucidating circR-loop subtypes, verifying structural conservation, species specificity, and half-life. Moreover, developing risk assessment models based on circR-loop for various diseases and validating its efficacy and stability as an emerging biomarker through large-scale clinical trials is imperative [5].
In terms of therapeutics, circRNA exhibits high stability, unique folding, and low immunogenicity, sparking interest in circRNA-based treatments. Novel RNA aptamers and protein translation vectors have shown promise in both in vivo and in vitro settings. These therapeutic modalities may artificially modulate circR-loop to regulate gene expression, thereby influencing cellular behavior [8, 45]. By targeting specific binding sites presented by circR-loop, these therapies may regulate the structural stability of circR-loop, akin to competitive endogenous RNA (ceRNA) mechanisms. This potential underscores the transformation of circRNAs into effective tumor treatments. However, circRNA-based aptamer therapy remains conceptual, facing challenges in molecular optimization design and mechanistic elucidation (46).
With ongoing advancements in basic circR-loop research, the scope of research methods continues to expand and optimize. Simultaneously, platforms for in vitro transcription, cyclization, and delivery systems are maturing, auguring clearer prospects for the clinical medical transformation of circR-loop.


留言 (0)