
記住我

We purified the human ATAD5-RFC complexes by co-expressing in insect cells the N-terminal-truncated ATAD5 (aa 812–end) and the human RFC2–RFC5 subcomplex (Fig. 1b and Extended Data Fig. 1a). The N-terminal-truncated ATAD5 comprises a minimal core that is capable of unloading PCNA from DNA31. In the rest of the paper, we refer to the N-terminal-truncated ATAD5 as ATAD5. Purified ATAD5-RFC could unload PCNA and was approximately 100-fold more efficient than was canonical RFC in PCNA unloading (Extended Data Fig. 2a,c–f). Moreover, the non-hydrolyzable ATP analog AMPPNP supported the unloading activity of ATAD5-RFC (Extended Data Fig. 2d), indicating that ATP binding, but not ATP hydrolysis, is required for unloading activity, consistent with an earlier study using weakly hydrolyzable ATPγS16.
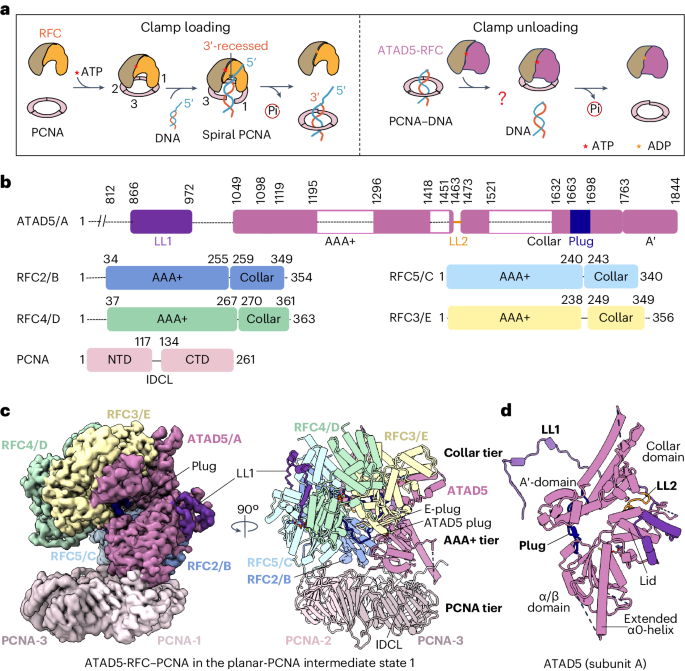
To assemble in vitro a complex of ATAD5-RFC bound to PCNA encircling dsDNA, we mixed purified ATAD5-RFC complex (Fig. 1b and Extended Data Fig. 1a) with PCNA that was preassembled with a 38-base-pair (bp) dsDNA in the presence of 1 mM ATPγS, a slowly hydrolyzable ATP analog, and incubated the mixture for 20 min. The PCNA-encircled DNA complex was preassembled following an established procedure38, which yielded around 60% PCNA–DNA complex in the mixture (Extended Data Fig. 1b). We then prepared cryo-EM grids and recorded a cryo-EM dataset. Two-dimensional (2D) and three-dimensional (3D) classifications resulted in an EM map at an overall resolution of 3.0 Å (Fig. 1c, Extended Data Figs. 1c,d, 3 and 4 and Table 1). The high-quality EM map allowed us to build an atomic model for most regions of ATAD5-RFC–PCNA, except for several disordered loops (Fig. 1b,d). AlphaFold-Multimer was used to predict the long N-terminal loop in ATAD5. The predicted model had a high confidence level in most loop regions and was consistent with the final atomic model (Extended Data Fig. 5a,b).
Table 1 Cryo-EM data collection, refinement and validation statisticsIn the ATAD5-RFC–PCNA complex, ATAD5-RFC sits above the PCNA ring, forming a three-tiered structure: ATAD5-RFC comprises the top collar tier and middle AAA+ tier, and the PCNA ring forms the bottom tier (Fig. 1d). The collar and AAA+ tiers are formed by the respective collar domains and AAA+ modules of ATAD5 and RFC2–RFC5 subunits arranged in a right-handed spiral. Because human RFC and yeast RFC were purified before structure determination and gene sequencing, the convention of numbering complex subunits by their respective size in a polyacrylamide gel resulted in the small RFC subunits having a different numbering scheme between yeast and human RFC. Thus, we also use the uniform convention of referring to clamp-loader subunits alphabetically, A, B, C, D and E, arranged counterclockwise when viewed from the carboxy-terminal collar domains. Thus, for human and yeast RFC, the subunits are A (hRFC1, yRfc1), B (hRFC2, yRfc4), C (hRFC5, yRfc3), D (hRFC4, yRfc2) and E (hRFC3, yRfc5) (Extended Data Fig. 6a). For ATAD5-RFC, the A subunit is ATAD5, which replaces RFC1. We found that four subunits (ATAD5, RFC2, RFC5 and RFC4, that is, subunits A–D) each bound an ATPγS, and RFC3 (subunit E) bound ADP (Extended Data Fig. 6b–f). The overall architecture and the nucleotide-binding pattern are shared with all reported clamp–clamp loader complex structures in the absence of DNA—from T4 phage to Escherichia coli, yeast and human15,19,20,21,39,40, and are similar to the 9-1-1 clamp loaders RAD17-RFC and Rad24-RFC25,26,27.
The middle AAA+ tier also contains the C-terminal A′ domain of ATAD5, which packs against the RFC3 AAA+ and forms the A-gate with the ATAD5 AAA+ module18 (Fig. 1d). The A-gate opens to admit the 3′-ss/ds DNA junction in a typical clamp loader, such as RFC19,20,21. We found that ATAD5 contains three unexpected features that are absent in any known clamp loader: an extended N-terminal loop, here called locking loop 1 (LL1), a second locking loop (LL2) and a plug (Fig. 1d); these elements seem to be crucial for ATAD5’s unloading function and will be discussed below. The structure is devoid of DNA, suggesting that, in our reaction, PCNA unloading by ATAD5-RFC was complete upon 20-min incubation, and PCNA returned to the closed planar ring in the complex (herein referred to as planar-PCNA intermediate state 1). However, although most PCNA rings (>60%) were bound to DNA before ATAD5-RFC was added to the mixture, it remains possible that ATAD5-RFC binds to PCNA after it slides off DNA. Indeed, we obtained a similar 3D structure to planar-PCNA intermediate state 1 by directly incubating ATAD5-RFC with free PCNA (Extended Data Fig. 7).
Two unique ATAD5-RFC locking loops keep the A-gate shutFor a PCNA loader, A-gate opening is a crucial step that results in opening of the PCNA ring, allowing DNA access to both PCNA and the central chamber of the loader19,20,21. As mentioned above, ATAD5 contains three unique features that are not present in human RFC1 (Fig. 1d). The first unique feature is LL1, which is 105 residues long (Gln868 to Ala1072) and wraps around the outer surface of the unloader complex, meandering around ATAD5 (subunit A), RFC2 (subunit B) and RFC5 (subunit C). It then inserts into a cleft between RFC5 (subunit C) and RFC4 (subunit D) (Fig. 2a). LL1 was well resolved in the EM map, and could be divided into four regions (Fig. 2a). Local interactions convert several LL1 regions into α-helices and β-strands. Indeed, LL1 adopts essentially the same conformation as the other conformers (which are discussed later). An earlier study showed that substitution of four residues in LL1 (872–875 aa) with four alanines severely decreased ATAD5-RFC unloading action in cells, consistent with the importance of LL1 for ATAD5-RFC function31. The second unique feature is that the alternative linker (AL) loop between the collar and the AAA+ module of ATAD5, which is conserved among the RFC loaders and unloaders, is positioned right above the A-gate and near LL1, suggesting that the AL has a role in locking the A-gate shut. We therefore renamed the AL loop in ATAD5 to locking loop 2 (LL2). LL2 forms three H-bonds with the RFC2 α/β subdomain: Asn1467 with RFC2 Thr191, the main chain oxygen of Asn1467 with RFC2 Thr56, and the main chain oxygen of Leu1462 with RFC2 Arg189. These H-bonds stabilize LL2 firming its apparent action as a lock that blocks the ATAD5 A-gate opening (Fig. 2b). In comparison, the RFC1 AL is flexible and enables A-gate opening18,21. The third unique feature, to be discussed in detail below, is the ATAD5 plug, comprising 36 residues (Leu1663–Asp1698), that occupies the central chamber (Fig. 2a).
Fig. 2: The ATAD5 LL1 locks the A-gate and prevents conformational changes in ATAD5-RFC.
a, Top view of ATAD5-RFC. The bound nucleotide in each subunit is shown as sticks, and Mg2+ in spheres. The EM densities for LL1 and the plug are shown as transparent gray surfaces. LL1 is divided into numbered four regions, to better show detailed interactions in c-e. b, LL1 and LL2 lock the A-gate shut by stabilizing the ATAD5 lid and collar. The zoomed inset shows detailed interactions of LL2 with the RFC2 α/β fold. c-e, Close-up views of the first to third regions in LL1 interacting with RFC4 (c), RFC5 (b) and RFC2 (e). f, Close-up view of the fourth region of LL1 (the helix hairpin) interacting with the ATAD5 lid domain. Key interacting residues are shown as sticks and labeled. H-bonds are shown as dashed lines.
The LL1 region 1 folds into a β-strand to complement the five-stranded β-sheet core (β-1–β-5) of the RFC4 α/β subdomain (Fig. 2c). Thus, this LL1 region is tightly coupled to the RFC4 α/β fold, and they can move together as a rigid body. The LL1 β-strand is parallel to the adjacent β5 and forms two additional H-bonds with the β1-strand: LL1 Gln868 with β1 Gln73, and LL1 His872 with β1 Tyr77. The LL1 region 2 is hydrophobic; it contains two short α-helices and wraps around the side of the RFC5 collar domain, in which LL1 residues Leu882, Leu885, Leu892 and Leu893 form hydrophobic interactions with the RFC5 collar residues Leu269, Leu272, Leu251, Leu255 and Phe293 (Fig. 2d). The LL1 region 3 is inserted into a cleft between the lid and collar domains of RFC2 and interacts primarily with the RFC2 lid (Fig. 2e): LL1 Leu919 forms a hydrophobic interaction with the RFC2 lid residues Leu199 and Ile222, LL1 Phe916 forms a π–π interaction with the RFC2 lid residues Phe223 and Ser907 and LL1 Ser917 forms an H-bond with the RFC2 lid residues Glu249 and Glu219. The LL1 region 4 contains a helix hairpin that, together with LL2, sandwiches the ATAD5 helical lid domain and holds the lid close to the RFC2 α/β subdomain, thereby preventing the A-gate from opening (Fig. 2b). Specifically, the helix hairpin of LL1 region 4 packs tightly against the ATAD5 lid domain, involving eight hydrophobic residues (Val943, Leu947, Leu948, Ile951, Phe958, Leu960, Phe964 and Leu967) (Fig. 2f). The lid α1-helix wedges into the hydrophobic cleft of the helix hairpin and contributes Leu1371 to the interaction. The LL1 region 4 also forms three H-bonds with the lid: Glu950 with lid Arg1376, Gln971 with lid Glu1373 and Arg936 with lid Asp1455. In summary, the extensive interactions stabilize the LL1, and in turn, the LL1 seems to tie the ATAD5-RFC into a stable structure that is resistant to the conformational changes that are required for binding of the same substrate (for example, ssDNA, dsDNA, gap, nick) as the clamp loaders in the central cavity18,19,20,21.
The ATAD5 plug prevents DNA from entering the ATAD5-RFCThe remaining key feature of ATAD5-RFC is the ATAD5 plug inside the central chamber (Figs. 1d and 2a). The RFC1 collar domain contains five α-helices (α1–α5) and connects to the A′-domain through a short loop (Fig. 3a,b). The ATAD5 collar domain also contains five α-helices, but α2 and α5 are much longer than the corresponding helices in human RFC1 (Fig. 3b). The ATAD5 plug is formed through extension of the linker loop between the α4 and α5 helices of the ATAD5 collar domain, as compared with the human RFC1 collar domain (Fig. 3a,b). The ATAD5 collar contains two additional insertions: a 110-residue insertion between α3 and α4 containing a short β-stand (β1) and disordered remaining region, and another insertion between α5 and A′-domain containing another short β-stand (β2) (Fig. 3a,b). The short β-stands of the two insertions form a parallel two-stranded β-sheet. The two inserted regions interact with the α2 helix of the lid: Tyr1757 and the main chain oxygen of Asn1755 form H-bonds with the lid residues Asp1378 and Lys1380, respectively (Fig. 3c). These insertions glue the lid and A′-domain together, and function together with LL1 and LL2 to block the A-gate (Fig. 3c).
Fig. 3: The unique ATAD5 plug occupies the central chamber of the unloader complex.
a, Side-by-side comparison of human ATAD5 and RFC1 (PDB 6VVO). The collar domain is colored, and all other regions are shown in light gray. The plug and associated insertion regions in the ATAD5 collar domain are colored blue and cyan, respectively. The disordered 112-aa connecting loop is shown by a dashed cyan line. b, Comparison of the collar domains of ATAD5 (left) and RFC1 (right). The conserved α-helices in both collar domains are labeled. Bottom: the ATAD5 plug is shown as sticks and is superimposed with the EM density in semitransparent gray surface view. c, Close-up of the cyan box in a, showing the ATAD5-plug-associated insertion region near the closed A-gate. The insertion region interacts with the ATAD5 lid domain to help LL1 and LL2 lock the A-gate. Key residues in the interaction are shown as sticks and labeled. d,e, Two close-up views of the blue box in a, in the context of the ATAD5-RFC pentamer. The ATAD5 plug forms a short antiparallel β-sheet with the conserved E-plug (d) and is stabilized by the AAA+ domains of RFC2, RFC3 and RFC5 (d,e). The ATAD5 plug also bridges the α/β fold and the A′ domain to rigidify ATAD5 (e). H-bonds are shown as dashed lines, and key interacting residues are shown in sticks and labeled. f, SDS–PAGE (8%) of 4 μg each of WT ATAD5-RFC and the five ATAD5-RFC mutants (Mut1–Mut5). Mut1–Mut4 contain either deletions or insertions in the truncated ATAD5 (Δ1–812). Mut5 has an amino acid replacement in the Arg finger residue of RFC3. The gel analysis was performed once. The label ‘MW stds’ refers to molecular weight standards. CT, C terminus. g, Comparison of unloading activity of WT ATAD5-RFC with ATAD5-RFC mutants; 6.5 nM ATAD5-RFC (either WT or mutants) was incubated with 32P-labeled PCNA–DNA and 2 mM ATP for 5 min at 37 °C, followed by gel filtration to determine whether the 32P-labeled PCNA is unloaded from DNA. The x axis label ‘Fraction’ refers to individual gel filtration fractions collected for activity assay. This experiment was performed once.
The ATAD5 plug contains a short β-strand that complements the two-strand E-plug (RFC3-Plug) (Fig. 3d). The E-plug is a conserved feature among all clamp loaders and unloaders: its two positively charged residues insert into the DNA main groove in all known clamp-loader structures15. However, in the ATAD5-RFC unloader, ATAD5 plug residues Gln1691 and Tyr1672 form H-bonds with E-plug residues Lys79 and Lys80, respectively, thereby neutralizing the E-plug’s ability to interact with DNA. The ATAD5 plug residues Arg1694 and Ser1693 form two additional H-bonds with Asp99 and the main chain oxygen of Pro77 in the E-plug, respectively (Fig. 3d).
The ATAD5 plug interacts with all ATAD5-RFC subunits, except for RFC4 (Figs. 1d and 3d,e). The ATAD5 plug residues Ser1686 and Asp1690 and the main chain oxygen of Leu1694 form H-bonds with Asp148 and Thr147 in RFC2 and Arg96 in RFC5, respectively (Fig. 3e). The ATAD5 plug also bridges the ATAD5 α/β domain and the A′-domains to rigidify the entire ATAD5 molecule. Specifically, Leu1688 in the ATAD5 plug interacts hydrophobically with Val1309 and Phe1311 in the α/β domain; Thr1685 forms an H-bond with Asp1338 in the α/β domain; Trp1679 inserts into a hydrophobic pocket in the A′-domain lined by Ile1772, Ile1775 and Ile1794; and Thr1680 forms an H-bond with the A′ domain Arg1790 (Fig. 3e). Therefore, the ATAD5 plug resides inside the central chamber and forms a number of interactions that rigidify the clamp-loader complex and fills its central chamber to the exclusion of DNA.
To functionally validate the unique structural features in ATAD5, we generated five ATAD5-RFC mutants (Mut1–Mut5) and performed the PCNA-unloading assay. In Mut1 (Δ1186–1294), the long flexible loop region of ATAD5 and part of the interface with PCNA was deleted; in Mut2 (Δ812–889), the LL1 was deleted; in Mut3, four residues (GGGS) were inserted in LL2; in Mut4 (Δ1668–1698), the A plug was deleted; and Mut5 had a p.R178A substitution in the RFC3 arginine finger region that inactivated the RFC4 ATPase site. Mut1 and Mut5 formed a stable ATAD5-RFC complex, and their PCNA-unloading activity was indistinguishable from that of the wild-type (WT) protein (Fig. 3f,g). However, Mut2, Mut3 and Mut4 were either not expressed well or were not stably incorporated into the RFC complex, indicating that LL1, LL2 and the plug have important roles in ATAD5-RFC structural integrity. Mut4 exhibited reduced assembly of the full complex than did the WT protein, with a corresponding reduction in unloading activity (Fig. 3f,g). This could simply reflect the lower concentration of full complex, considering that a reduction of WT ATAD5-RFC concentration from 10 nM to 2.5 nM led to a similar decrease in unloading efficiency (Extended Data Fig. 2e). Therefore, we cannot rule out the possibility that Mut4 is as active in unloading as the WT protein. In conclusion, owing to a loss of structural integrity of Mut2, Mut3 and Mut4, it remains unclear whether LL1, LL2 and the plug have a direct role in PCNA unloading.
ATAD5 has a larger interface with PCNA than RFC1Three subunits of ATAD5-RFC interact with two of the three PCNA subunits: ATAD5 (subunit A) and RFC2 (subunit B) contact PCNA-1, and RFC5 (subunit C) contacts PCNA-2 (Fig. 1c). Notably, no subunit contacts PCNA-3. The PCNA-binding interface of ATAD5 (subunit A) is larger than those of RFC2 and RFC5 (subunits B and C) (Fig. 1c). These interaction patterns resemble those in several recently determined RFC–PCNA structures in an autoinhibited state that occurs before RFC opens the PCNA ring for loading onto DNA15,19,20. However, ATAD5-RFC is a clamp-unloading complex and needs to first interact with the DNA-bound PCNA. Therefore, we propose that the closed planar-PCNA state represents a late intermediate stage in the unloading process in which ATAD5-RFC has unloaded the PCNA from DNA, and the PCNA ring has closed and is ready to dissociate from ATAD5-RFC, as explained later.
A unique feature in ATAD5 is the much-extended α0-helix in the AAA+ domain, compared with that of human RFC1 (PDB: 6VVO)15 (Fig. 4a,b). The ATAD5 α0-helix is long enough to reach PCNA and interact with the PCNA-1 C terminus through three H-bonds between Gln1093 in α0 and PCNA Glu258, Gln1094 in α0 and the main chain oxygen of Glu256 in PCNA, and Lys1098 in α0 and the main chain oxygen of Gln204 in PCNA-1 (Fig. 4c). Furthermore, the α2 element of ATAD5 contains a non-consensus PCNA-interacting peptide (PIP) motif (1173-EATQSHQV-1180). The PIP motif inserts into the PIP pocket and forms four H-bonds with PCNA-1: the main chain oxygen of Gln1170 in the PIP motif with Ser43 in PCNA-1; Gln-1173 in the PIP motif with Lys254 in PCNA-1; the main chain nitrogen of His1178 in the PIP motif with the main chain oxygen of Pro253 in PCNA-1; and the main chain nitrogen of Val1180 in the PIP motif with the main chain oxygen of His44 in PCNA-1. Another unique feature in ATAD5 is an extra β-strand (β3) in the AAA+ domain following the PIP motif (Fig. 4d). ATAD5 β3 interacts with PCNA-1 through an additional H-bond between Gln1188 in the β3-strand and Asp122 in PCNA-1. These two unique features resulted in a large interface between ATAD5 and PCNA-1, spanning 1,295 Å2. In comparison, human RFC1 contains the classic PIP motif that inserts into the PIP pocket of PCNA-1, and forms hydrophobic interactions with the Phe702 and Tyr703 residues of PCNA-1 (ref. 15). Human RFC1 also forms three H-bonds with PCNA: Asn695 in RFC1 with Lys254 in PCNA-1; Thr697 in RFC1 with the main chain oxygen of Lys254 in PCNA-1; and Ser698 with the main chain oxygen of His44 in PCNA-1 (Fig. 4e). The interface between human RFC1 and PCNA-1 is 1,009 Å2. The significantly larger PCNA-binding interface between ATAD5 and PCNA (1,295 Å2) could contribute to ATAD5’s unloading activity. Finally, the ATAD5 AAA+ domain contains an unusually long 100-residue loop connecting the β3- and β4-strands. This loop is disordered in our structure, but it can’t thread through the small tunnel between the ATAD5 AAA+ domain and RFC2 (subunit B) and must wrap around the ATAD5 α/β subdomain, thereby passing through the central chamber of the ATAD5-RFC complex (Fig. 4a). This long loop could contact the inner face of the PCNA ring. However, deleting the flexible loop in the ATAD5 AAA+ domain (Mut1) did not markedly affect the PCNA-unloading activity by the unloader (Fig. 3f,g). Therefore, the function of the flexible loop is unclear.
Fig. 4: The AAA+ module of ATAD5 contains a PCNA-binding interface larger than that of RFC1.
a, The interface between ATAD5 and PCNA-1. The ATAD5 non-canonical PIP box is colored yellow, and the two additional PCNA-binding regions are colored mint green and blue. The secondary structures in the α/β fold are labeled. b, The interface between human RFC1 and PCNA-1 (PDB: 6VVO). RFC1 binds PCNA-1 by only the conserved PIP box (yellow). c, Close-up view of the interactions between the ATAD5 α0 and PCNA-1 (dashed green square in a). d, Close-up view of interactions of the ATAD5 PIP box and the extended β3-strand with PCNA-1 (dashed magenta square in a). e, Close-up view of the interactions of the conserved RFC1 PIP box with PCNA-1 (dashed yellow square in b). Residues involved in the interactions are shown in sticks and labeled in c-e.
ATP hydrolysis by RFC4 might open PCNAThe structure in Fig. 1 (planar-PCNA intermediate state 1) was obtained from a reaction that lasted for 20 min. To look for earlier intermediates in the reaction, we next used cryo-EM to examine three assembly mixtures incubated for decreasing amounts of time, ranging from 10 to 6 to 3 min, to visualize the intermediates during ATAD5-RFC unloading of PCNA from DNA. We did not observe any complex bound to DNA in these samples (Extended Data Fig. 1d). However, we obtained from the 3-min incubation mixture two other intermediate states (cracked-PCNA intermediate 2 and disordered-PCNA intermediate 3) of the ATAD5-RFC–PCNA binary complex, at average resolutions of 3.5 Å and 3.1 Å, respectively (Fig. 5a,b, Table 1 and Extended Data Figs. 8 and 9). ATAD5-RFC does not undergo large conformational changes in these intermediates; in particular, the conformations of the unique features (such as LL1, LL2 and the plug) of ATAD5 in these states are the same as in the planar-PCNA intermediate state 1 that was obtained from the 20-min incubation sample. But the PCNA rings in these intermediate states are different from the planar-PCNA intermediate state 1 (Figs. 1c and 5a,b). In the cracked-PCNA intermediate state 2, all three PCNA molecules are ordered and form a right-handed spiral with a crack between PCNA subunits 1 and 3—but not a gap (Fig. 5a). In the disordered-PCNA intermediate state 3, PCNA-1 and PCNA-2 are well resolved, but PCNA-3 is disordered and invisible (Fig. 5b). We performed 3D variability analysis (3DVA) and found that the PCNA-3 undergoes a large-scale movement (Supplementary Video 1): PCNA-3 tilts down to form a cracked interface with PCNA-2 on the left, then slides upward to form a new cracked interface with PCNA-1 on the right. From the disordered-PCNA intermediate state 3 dataset, we derived another intermediate structure (open-PCNA intermediate state 3′) at an average resolution of 4.2 Å (Extended Data Fig. 8) that contains a 5-Å gap between PCNA-2 and PCNA-3 (Fig. 5c). This gap is unique because, to date, all structures of clamp loaders with open PCNA have a gap between PCNA-1 and PCNA-3.
Fig. 5: ATP hydrolysis by RFC4 underlies the transition of the PCNA crack from the right to the left of PCNA-3.
a-c, EM maps (top; postprocessed with DeepEMhancer44) and atomic models (bottom) of ATAD5-RFC bound to PCNA in intermediate states 2 (a), 3 (b) and 3′ (c). Maps and structures are colored by individual subunits, as in Fig. 1b. d,e, Close-up views of the nucleotide-binding regions in RFC4 (subunit D) in intermediates 2 (d) and 3 (e), corresponding to areas marked by the blue and red squares at the bottom of a and b. The nucleotides and interacting residues are shown as sticks and labeled. f, The open (gapped) PCNA structure in the intermediate state 3′. The PCNA structure of intermediate state 2 is superimposed, but only the position of its PCNA-3 is shown (in cyan), to reveal a 30-Å movement of PCNA-3 between intermediate state 2 and intermediate state 3′.
Unexpectedly, the RFC4 (subunit D) in the disordered-PCNA and open-PCNA intermediate states 3 and 3′ is occupied by ADP. The ATPγS in RFC4 of cracked-PCNA intermediate 2 is stabilized by Val41, Lys84 and an Mg2+ coordinated by Thr85 and Asp150. By contrast, the ADP in RFC4 of states 3 and 3′ is stabilized by Val41, Thr85 and Arg239 (Fig. 5d,e). Therefore, the ATPγS in RFC4 seems to have been either hydrolyzed to ADP or exchanged for ADP in the transition between the cracked-PCNA and disordered-PCNA intermediate states 2 and 3 (Fig. 5c,d and Extended Data Fig. 10). Superimposition of PCNA-1 in the cracked-PCNA and open-PCNA states shows that PCNA-3 moves by 30 Å (Fig. 5f). We suggest that the cracked versus open PCNA state is due to different nucleotide occupancies in these ATAD5-RFC intermediates.
What might these intermediate states suggest about the role of ATP hydrolysis? Although this report does not claim to describe the unloading mechanism of ATAD5-RFC, it sheds light on a possible function of hydrolysis. Consider the canonical RFC. RFC needs only to bind ATP to open or close PCNA around DNA; ATP hydrolysis is needed for RFC to eject from PCNA encircling DNA18. This is similar to ATAD5-RFC, which needs only to bind ATP to open PCNA for unloading (Extended Data Fig. 2d). Consistently, inactivation of RFC4 ATPase (Mut5, Arg finger mutant RFC3-R178A) did not affect PCNA-unloading activity (Fig. 3g). We therefore presume that, similar to RFC, ATAD5-RFC uses ATP hydrolysis to dissociate from PCNA, allowing the complex to be reused in the unloading of other PCNA clamps on DNA. This proposed action predicts that PCNA may alter the ATPase activity of ATAD5-RFC. Indeed, we found that PCNA stimulated the ATPase activity of ATAD5-RFC by fivefold (Extended Data Fig. 2b). However, DNA had no effect on the ATPase activity of ATAD5-RFC (Extended Data Fig. 2b), unlike RFC which is stimulated by DNA. These ATPase properties are consistent with those of yeast Elg1-RFC23.
The RFC5 AAA+ module may control PCNA ring closureComparing the closed-PCNA and cracked-PCNA intermediate states 1 and 2 reveals that both PCNA-2 and PCNA-3 undergo a large conformational change, in which PCNA-2 and PCNA-3 tilt by 10° and 20°, and their N-terminal halves move by 14 Å and 24 Å, respectively, with PCNA-1 remaining stationary (Fig. 6a,b). The planar-PCNA and cracked-PCNA intermediate states 1 and 2 were captured by incubating the reaction mixtures of ATAD5-RFC and PCNA–DNA for 20 min or 3 min, respectively. Their corresponding subunits are similar, with a small r.m.s. deviation (r.m.s.d.) (~0.6 Å) for ATAD5, RFC2, RFC4 and RFC3, and a much larger r.m.s.d. (1.2 Å) for RFC5. Superimposition of the two RFC5 structures shows that the α/β fold domain and the helical lid domain are orthogonal (90°) in the cracked-PCNA intermediate state 2 but are 79° apart in the planar-PCNA intermediate 1 (Fig. 6c). In the cracked-PCNA intermediate state 2, RFC5 interacts with PCNA-2 through three elements, leading to an extensive interface of 3,044 Å2 (Fig. 6d): the non-canonical PIP motif (Ser110 to Lys117), the α1-extension-helix and the N-terminal peptide (Met1 to Ala13). The RFC5 PIP residues Phe115 and Ile114 are inserted into the PCNA-2 PIP-binding cleft, lined by Leu47, Leu126 and Ile128. The RFC5 PIP motif also forms several H-bonds with PCNA-2: Ser110, Arg112 and Lys117 in the PIP motif form H-bonds with Lys254 and the main chain oxygens of Ala231 and Asp232 in PCNA-2. The α1-extension-helix forms two H-bonds with PCNA-2, between the main chain oxygen of Gly84 and Asp80 in RFC5 and His44 and Glu124 in PCNA-2, respectively. The RFC5 N-terminal α-helix forms two H-bonds with the PCNA-2 interdomain connector loop, between Gln8 and Gln9 in RFC5 and Asp122 and Asp120 in PCNA-2, respectively. In the planar-PCNA intermediate state 1, the interface is smaller (2,059 Å2; Fig. 6e): the aromatic residue Phe115 of the PIP box also inserts into the PIP-box binding cleft of PCNA-2, and Arg112 in the PIP motif forms a single H-bond with the main chain oxygen of Lys254 in PCNA-2. The RFC5 N-terminal α-helix and the α1-extension-helix move up and lose contact with PCNA-2. Additionally, RFC4 interacts with the PCNA-3 C-terminal loop through an H-bond between Gly138 in RFC5 and Ser261 in PCNA-3 in the cracked-PCNA intermediate 2 (Fig. 6f). Owing to the numerous RFC5–PCNA interactions, we suggest that movements of the RFC5 AAA+ module control PCNA, converting it between a cracked spiral and a planar ring.
Fig. 6: Conformational change in RFC5 underlies a lock-washer to planar-ring transition of PCNA.
a, Superimposition of ATAD5-RFC–PCNA in intermediate states 1 and 2. Intermediate state 2 is shown in color, and state 1 in gray. b, Two side views of the superimposed PCNA structures showing that PCNA-2 and PCNA-3 undergo large rigid-body movements from a right-handed spiral to a planar ring. c, Overlay of RFC5 (subunit C) in intermediate state 2 (cyan) and in state 1 (gray). The α/β domain rotates 11° with respect to the helical lid. d–f, Close-up views of the three boxed regions in a, showing detailed interactions between the RFC5 PIP motif and PCNA-2 in intermediate state 2 (d), between the RFC5 PIP motif and PCNA-2 in intermediate state 1 (e) and between RFC4 and PCNA-3 in intermediate state 2 (f). The red box in d is enlarged on the right to show the RFC5 N terminus interacting with PCNA-2. Key residues are shown as sticks and labeled.
ATAD5-RFC opens a different gap in PCNA than RFCIn the planar-PCNA intermediate state 1, the interactions involving PCNA-1 remain the same, but the N terminus of RFC5 and the shorter α-helix lose contact with PCNA-2, and no subunit is in contact with PCNA-3 (Fig. 7a). In the cracked-PCNA intermediate state 2, ATAD5 contacts the middle of PCNA-1, RFC2 (subunit B) contacts the interface between PCNA-1 and PCNA-2, RFC5 (subunit C) contacts PCNA-2, and an RFC4 (subunit D) loop weakly binds PCNA-3 (Fig. 7b). In the open-PCNA intermediate state 3′, there is a 5-Å gap between PCNA-2 and PCNA-3 (the PCNA-3/2 gap). PCNA homotrimer exhibits a spiral shape in both the cracked-PCNA and open-PCNA states 2 and 3′, but only in the open-PCNA intermediate 3′ is PCNA-3 free to move further down to enlarge the gap (Fig. 7c). There is no PCNA-3/1 gap in all three observed ATAD5-RFC intermediate states.
Fig. 7: The PCNA ring is opened by ATAD5-RFC and by RFC at different locations.
留言 (0)