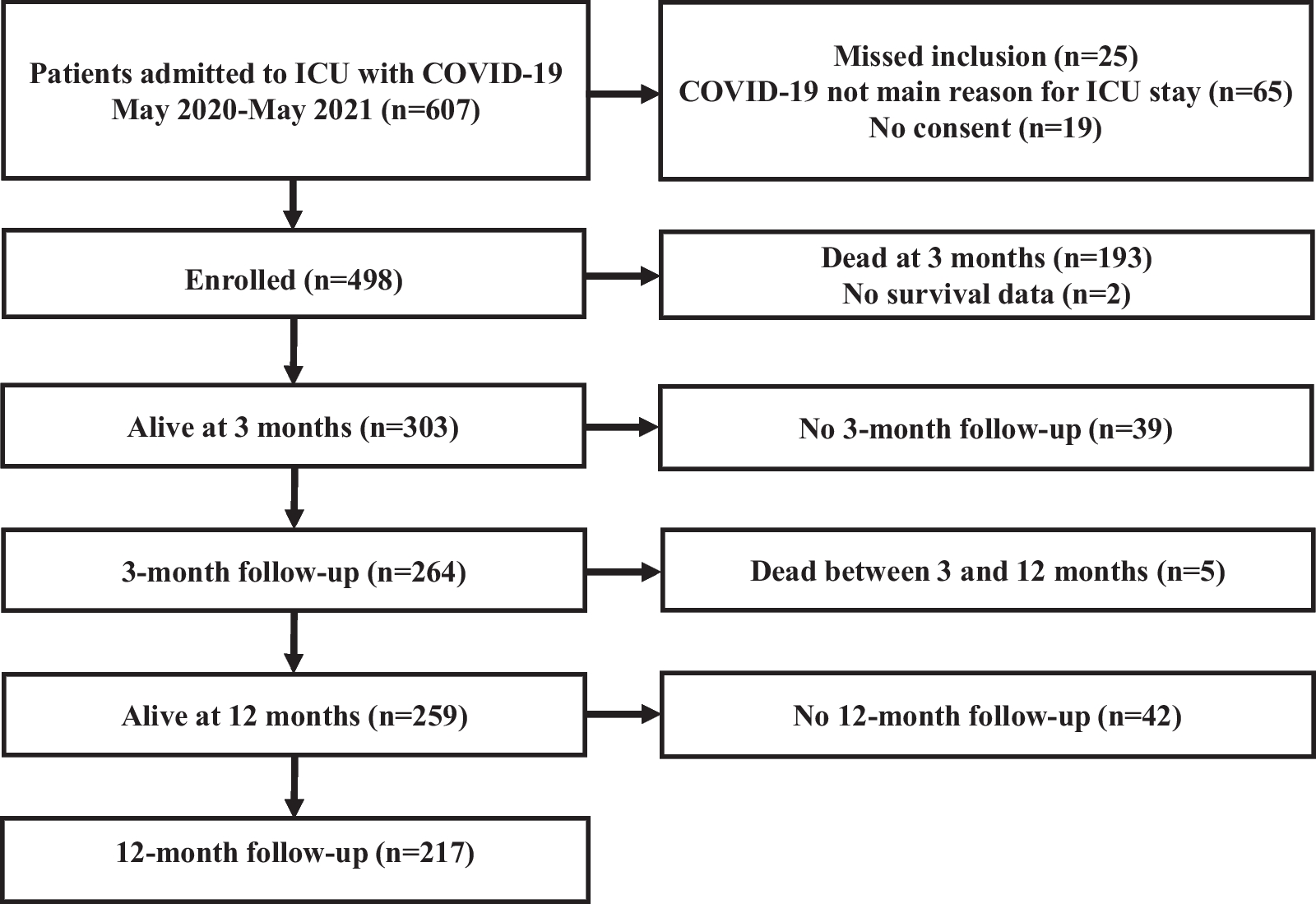
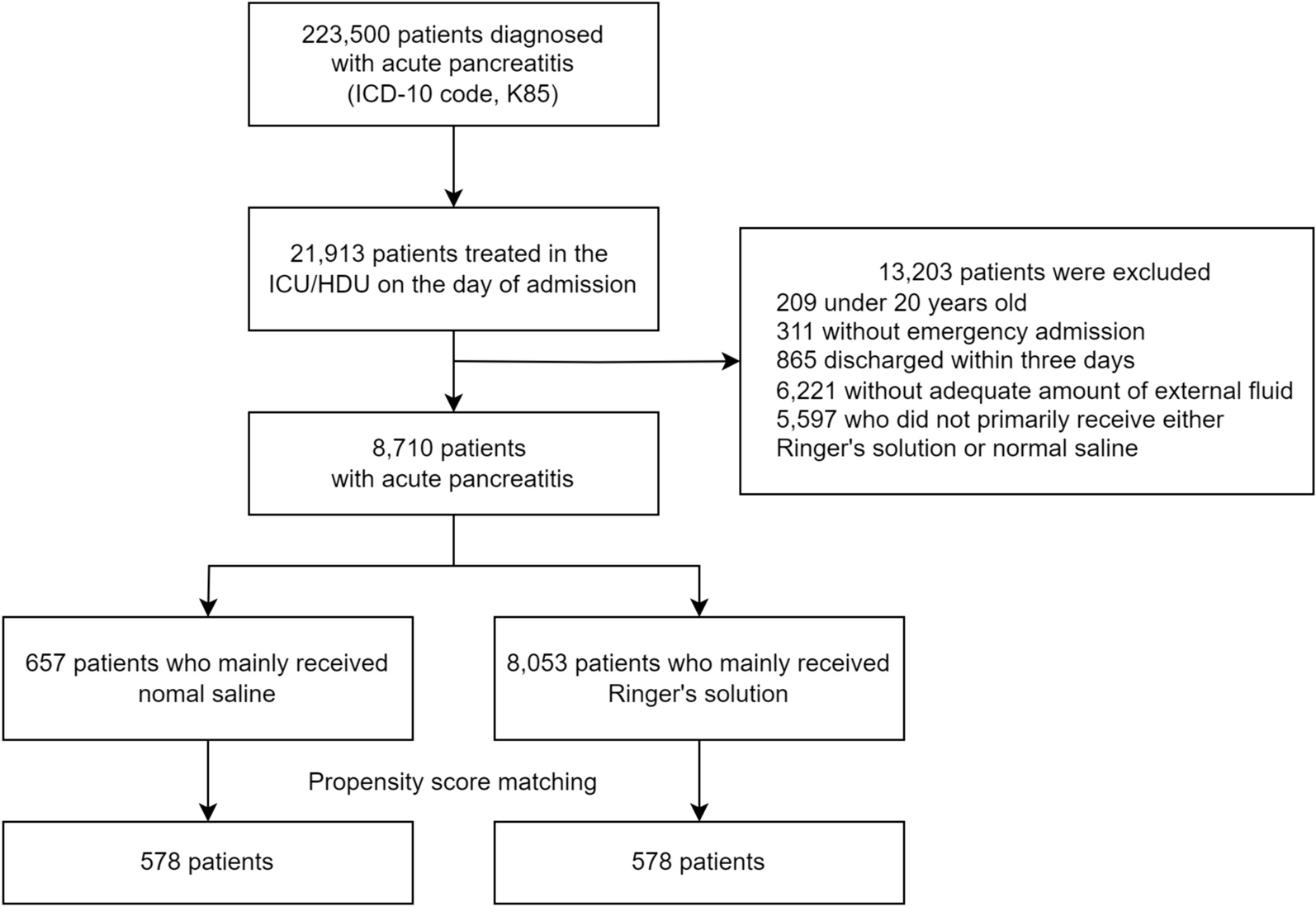
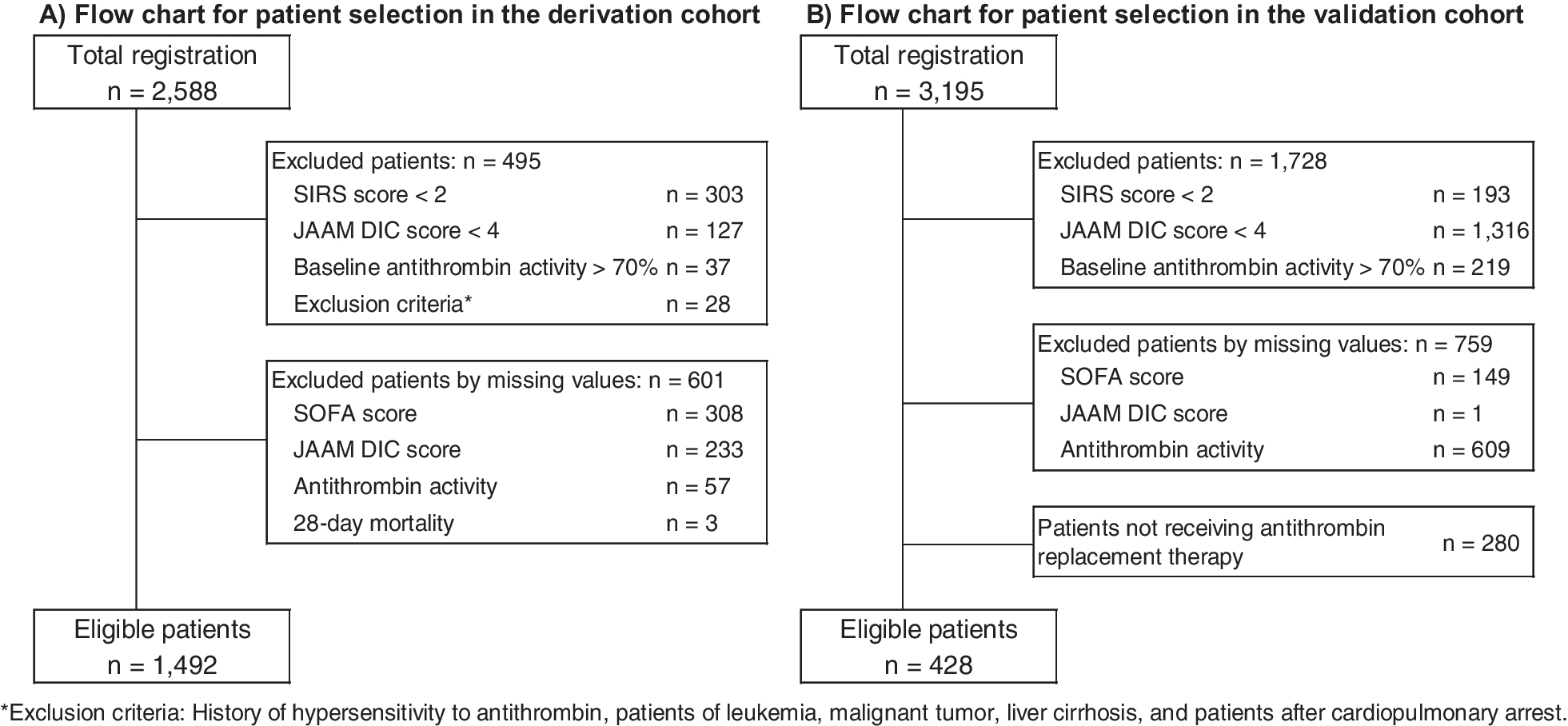
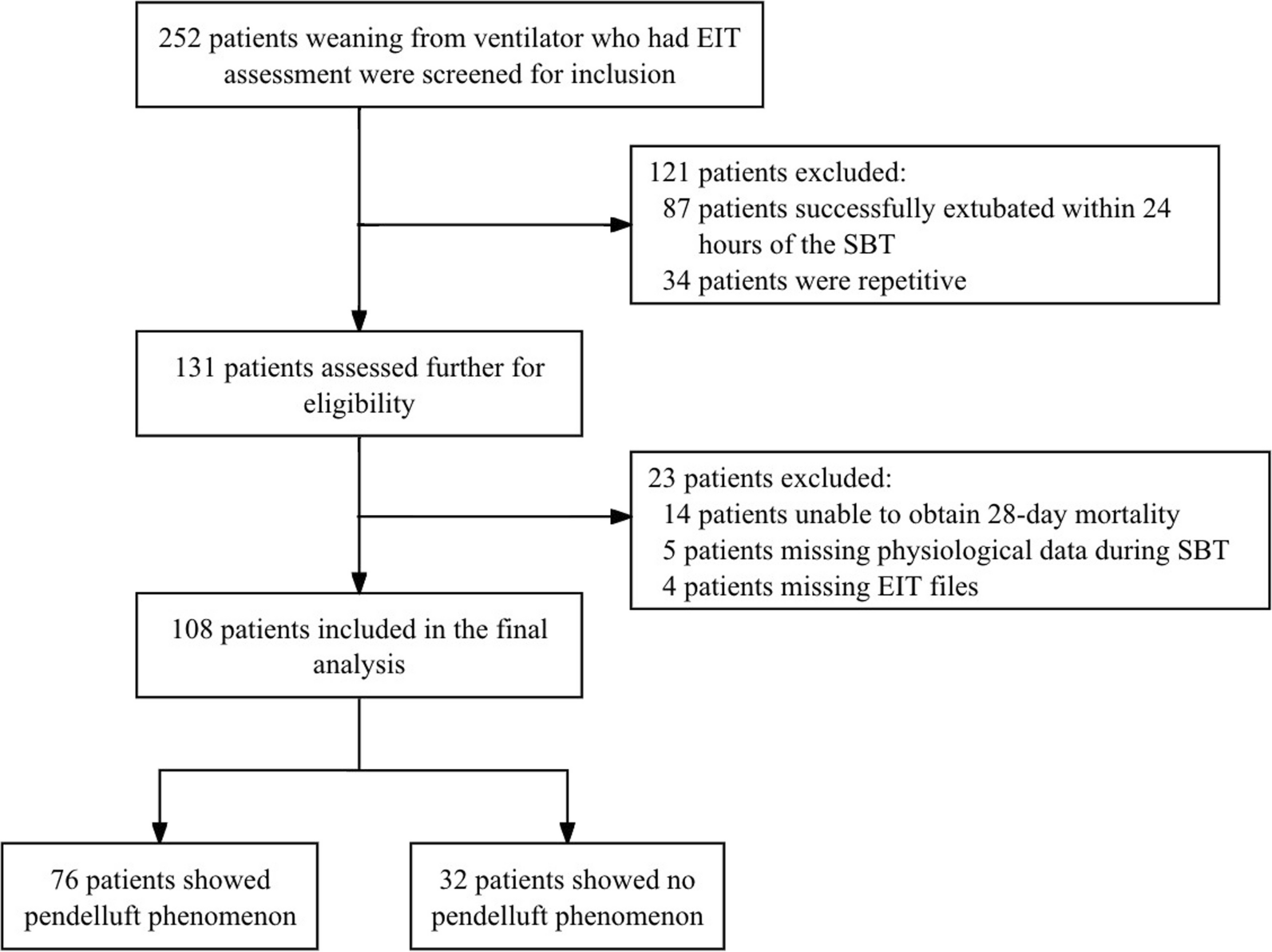
Main results
This post hoc study analyses the effects on the right ventricle and pulmonary vasculature of changing preload and afterload during VA ECMO weaning. Changing ECMO flow was used to model changing preload [21], such that increases in QECMO reduced pulmonary blood flow, thereby unloading the right ventricle. These changes led to decreases in right and left atrial pressures, as blood volume was redistributed from the pulmonary to the systemic compartment [22, 23], and increased total blood flow (i.e., increased venous return) [21]. The decreasing left atrial pressures indicate that the left ventricle coped well with the afterload produced by the VA ECMO [2]. During QECMO reductions, mean PA pressure increased as a consequence of increasing pulmonary blood flow [24], with a simultaneous decrease in input resistance (Z0) and characteristic impedance (Zc), attributable to improved distensibility and recruitment of the pulmonary vasculature [16, 25].
Effects of afterload increase
HPV and PE conditions acutely increased RV afterload. These changes were accompanied by small increases in RAP, indicating that the ventricles did not fail [26]. Rather, they operated below their stressed volumes [9, 26]. Both HPV and PE caused major increases in Z0.Zc remained unchanged compared to baseline as previously described [19, 20, 27].
During HPV and PE, the RV coped with acute afterload increases by increasing contractility (increased Ees) to maintain QLung when QECMO was decreased. This is in line with homeometric adaption (RV Anrep effect) [12, 17, 28]. Increased contractility from homeometric adaptation manifests as increased ventricular elastance (Ees), to match the increases in afterload (i.e., arterial elastance, EaExp). This mechanism restores RV–PA coupling to normal values (approximately 1.8 to 2.0; Figs. 2 and 5) [29] and establishes optimal energy transfer from the RV to the pulmonary vasculature [12]. Increases in ventricular elastance, in turn, lead to an increase in total (WT) and oscillatory hydraulic work (WOsc) with a constant WT/ WOsc ratio.
Effects of the respiratory cycle
When preload was reduced by increasing QECMO, the RV became more susceptible to afterload increases through the respiratory cycle, demonstrated by increases in EaInsp during baseline and PE conditions. The resulting inspiratory RV–PA decoupling (Figs. 2 and 5) lead to reduced SV and enhanced SVV (Fig. 3B). This resulted from a combination of decreased filling and acute RV ejection inability. During states of normal preload (i.e., low QECMO), the main cause of SVV was decreased filling—as demonstrated by only minor changes in RV–PA coupling. The additional relative respiratory cycle change in SV during conditions of low preload (i.e., high QECMO) was a result of an acute RV inability to eject against the increased afterload (increased intrathoracic pressure). Previous studies explain inspiratory SV decrease predominantly as an effect of increased afterload [30, 31]. The time course of homeometric RV adaptation (Anrep effect) is unknown [12]. Respiratory cyclic changes of afterload occur too rapidly to allow for homeometric adaptation or “slow response” [32]. At low preload, the Starling mechanism (heterometric adaption), may not suffice to increase SV during inspiration, and does not restore ventricular–arterial coupling to preserve ejection fraction.
The increase in RV afterload was highest during HPV, as demonstrated by the highest input resistance (Z0), mPAP, and EaExp. Porcine pulmonary vasculature is highly reactive [20, 33]. Nevertheless, changes in inspiratory load at HPV condition were less pronounced compared to PE or baseline since only one lung was exposed to positive pressure ventilation.
Right ventricular behavior
Our study group has demonstrated that during VA ECMO, cardiac output can be estimated using gas exchange or modified thermodilution and that RV ejection fraction can be assessed from the exponential decay of the thermodilution signal [5, 6, 16, 34, 35]. In line with the findings of the present analysis, we could demonstrate that the EDV/ESV relationship was linear [16, 36]. The slope of EDV/ESV represents 1 – RVEF. We can now extrapolate that the expiratory RVEF, remained constant during QECMO variations. This suggests that the RV dilated to cope with increased preload [26], shifting the PV loop rightwards (Fig. 2). This dilation caused almost no increase in RAP. The RV end-diastolic PV relationship is flat at normal operating volumes [26]; increases in RAP by additional volume loading would therefore indicate RV failure [37]. During inspiration, the brisk combination of decreased preload and increased afterload prevented the ventricle from acute dilation and adaption, explaining the inspiratory decoupling and decrease in ejection fraction [30]. The PV loop is shifted left- and upwards (Fig. 2). While expiratory values remain constant and support the previous data on EDV/ESV relationship [16, 26, 36], we could demonstrate that acute inspiratory increases in afterload (e.g., EaInsp) inhibit adequate RV ejection. This was most pronounced during states of low preload. Vieillard-Baron and colleagues had similar findings in echocardiographic studies: the respiratory cycle induced decreases in RV stroke volume, which were not associated with decreases in filling parameters and therefore interpreted as an afterload phenomenon [30, 31, 38]. It appears that although the RV can adapt to increases in afterload by increasing its contractility [28], the time available during the respiratory cycle is insufficient to allow for this adaptation [39]. The large variation in SV (Additional file 2: Fig. S1) may reflect RV–PA uncoupling rather than a pure preload dependency and is a result of preload decrease and simultaneous afterload increase [30, 40].
Clinical implications
Since the RV and PA are exposed to the same pressure at end-systole, ventricular–arterial coupling can be visualized as two intersecting lines at the end-systolic pressure (Fig. 2). This intersection divides the abscissa into end-systolic volume (ESV) on the side of Ees, and SV on the side of Ea. The RV and PA are exposed to the same pressure at end-systole. This end-systolic pressure provides a physiological and mathematical link between ventricular and arterial elastance, so that the ejection fraction can be determined by the Ees/Ea ratio or VA coupling [18]:
$$\frac=\frac}}}}+ 1 =\frac}}+ 1.$$
This is illustrated in Fig. 5 when comparing plots A&B with C&D [18]. From this equation, optimal coupling (ratio 1.5 to 2) will result in a RVEF of 60 to 67%.
When assessing right ventricular performance in a VA ECMO weaning trial, the EF, commonly accepted as a surrogate for contractility, depends on loading conditions and is an expression of RV–PA coupling [41]. It, therefore, seems reasonable to hypothesize that therapeutic measures improving contractility (for example, inotropes) and afterload (vasopressors or vasodilators, depending on the clinical context) should aim to improve coupling. In return, the right ventricular ejection fraction will increase. Pathological conditions leading to RV–PA decoupling will, therefore, inevitably lead to reduced ejection fraction, and with a clear understanding of coupling mechanisms, therapeutic options may be tailored to the underlying pathology. It is common to interpret a reduced ejection fraction as a consequence of low contractility. While this may be true in many cases, our results demonstrate that the ejection fraction is determined by contractility in combination with afterload and preload conditions. Lowering pulmonary vascular resistance (i.e., lowering pulmonary vascular elastance, Ea), optimizing volume status, or changing respiratory parameters and thereby decreasing right ventricular afterload may improve RV–PA coupling as efficiently as increasing contractility (Ees). We demonstrate that positive pressure ventilation in states of high afterload impacts right ventricular performance significantly, and positive pressure ventilation, therefore, may contribute significantly to disturbances in RV–PA coupling. If high pulmonary vascular resistance (i.e., high pulmonary vascular elastance, Ea) results from high left ventricular filling pressures, pharmacological or mechanical unloading of the left ventricular may optimize RV–PA coupling (and thereby RVEF) and interventions aimed directly at the right ventricle or pulmonary vasculature may not be necessary [2, 42]. It is therefore appealing to guide therapy and weaning from ECMO towards optimized coupling by directly influencing Ees (e.g., by increasing contractility) and Ea (by optimization of afterload and preload). Our analysis shows that right ventricular dilation is a physiological behavior of the right ventricle (i.e., heterometric adaption) to maintain RV–PA coupling. This should be considered in echocardiography guided weaning trials, where RV dilation is often seen as a weaning failure [43]. We demonstrate that dilation is a physiological adaptation if the RV operates below its stressed volume (i.e., at normal filling pressures). While we demonstrate the link between coupling and ejection fraction in the specific setting of VA ECMO, this physiological link, and the ensuing therapeutic targets (i.e., targeting Ea or Ees) could be applied to any hemodynamic assessment.
Limitations
We estimate higher RV ejection fractions in the current analysis than with our thermodilution approach. The reasons may be twofold: underestimation of the ejection fraction is common with thermodilution [44, 45]. We have shown that the exponential decay of thermodilution signals depends on the distance from the injection port [35]. Opposed to thermodilution studies where RVEF is estimated as a mean over multiple respiratory cycles, the elastance-based approach allows per beat calculation of EF during fractions of the respiratory cycle. Our findings are comparable to those of other groups [28].
We lack direct RV pressure signals. Instead, we rely on a modified single-beat method to estimate Pisomax, suggested by Pinsky [9]. Our estimations of Ea and Ees match published data from porcine PV loops [28]. We, therefore, judge the method to describe RV behavior reliably but conductance catheters are the gold standard to assess end-diastolic and end-systolic pressure–volume relationships [46].
The creation of HPV through main-stem intubation instead of using low levels of oxygen is a consequence of the retrospective nature of our study. In the protocol, we investigated the effects of shunt on a modified Fick principle. Therefore, both lungs were not evenly pressurized during HPV. Nevertheless, the diminished effect of lower intrathoracic pressure and one-lung ventilation on the reduction of RVEF and stroke volume variation highlights the impact of optimized ventilatory strategy in the setting of high right ventricular afterload.
Last, we lack transmural pressures in this experiment, which precludes us from fully differentiating the changes in stroke volume as a change of pre- or afterload [38, 46]. Additionally, our data did not allow for reconstruction of the end-diastolic PV relationship. The method of single-beat estimation from left ventricular EDPVR is not validated for RV use [47].



留言 (0)