
記住我
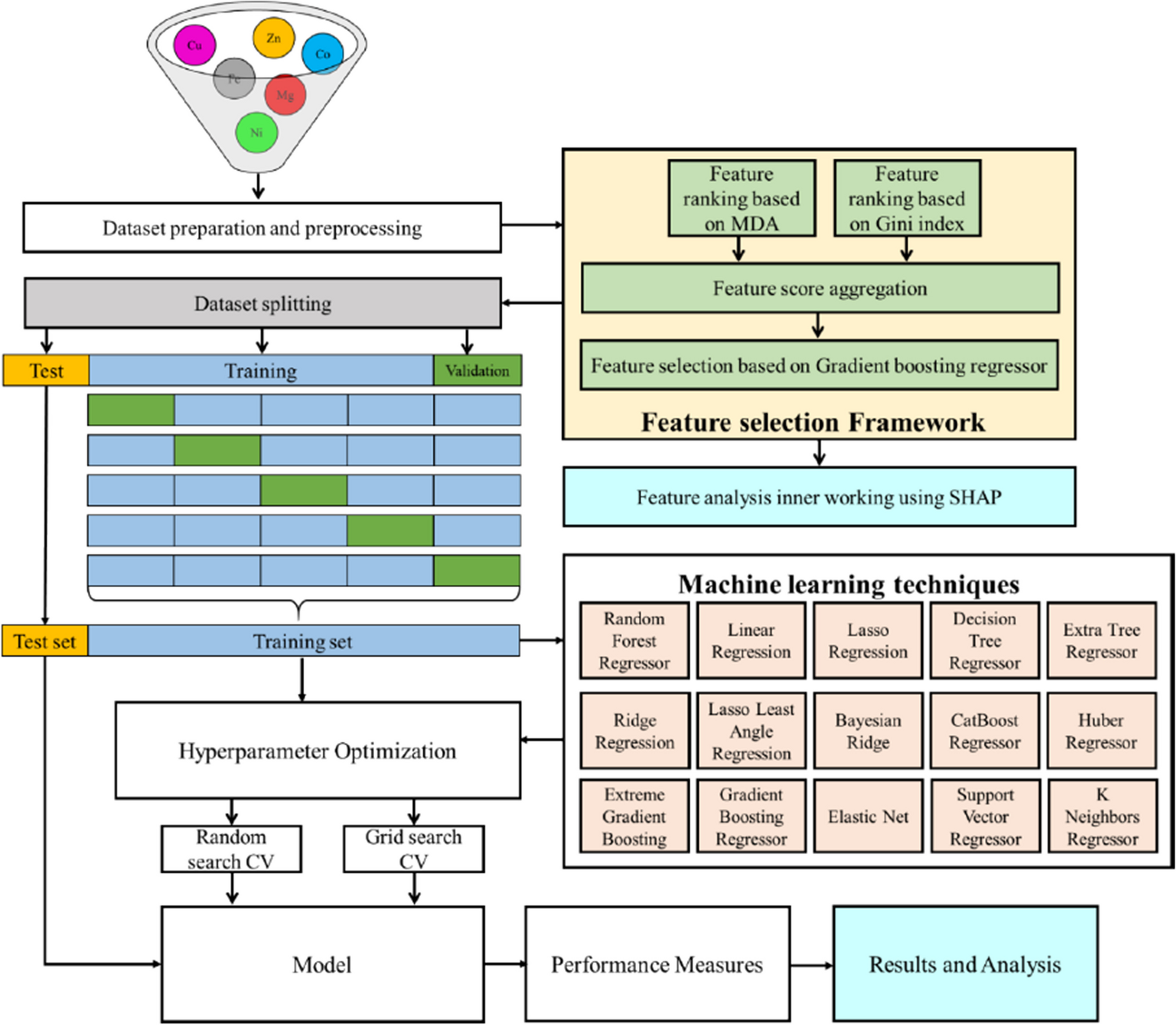

Thirty days after inoculation, the root systems of rough lemons inoculated with the SWK, GJ, FZ, GZ, YJ, and DBSR populations were weak and had evident brown spots (Fig. 1). Plant height, root length, and root weight of the six treatments were significantly lower than those of the control (p<0.05) and the number of nematodes in the root and rhizosphere soils was significantly higher than those in the control (p<0.05). However, the average nematode number in the roots of the XIN, ML, HN6, and HL inoculation treatments was only 1–2, and the nematode number in the rhizosphere soil was less than 10, which was not significantly different from that of the control (p>0.05). The root length difference between these four treatments was also not significant, and the root length and weight of XIN and HN6 inoculation treatments were not significantly different from those of the control (p>0.05) (Table 2). Therefore, SWK, GJ, FZ, GZ, YJ, and DBSR populations could parasitise rough lemons and had clear pathogenicity, whereas XIN, ML, HN6, and HL populations did not parasitise rough lemons.
Fig. 1
Symptoms of Citrus limon infected with 200 female nematodes of Radopholus similis per plant for 30 days. CK: non-inoculated rough lemon seedling and root system; XIN, ML, SWK, HN6, HL, GJ, FZ, GZ, YJ and DBSR: the rough lemon seedlings and root systems were inoculated with the different populations of R. similis, which originated from Zingiber officinale Roscoe (XIN), Crataegus pinnatifida (ML), Chrysalidocarpus lutescens (SWK), Musa AAA Giant Cavendish cv.Baxi (HN6), Maranta arundinacea (HL), Citrus reticulata (GJ), Anthurium andraeanum ‘Pink Champion’ (FZ), Anthurium andraeanum Linden (GZ), Curcuma longa (YJ), and Anubias nana (DBSR), respectively
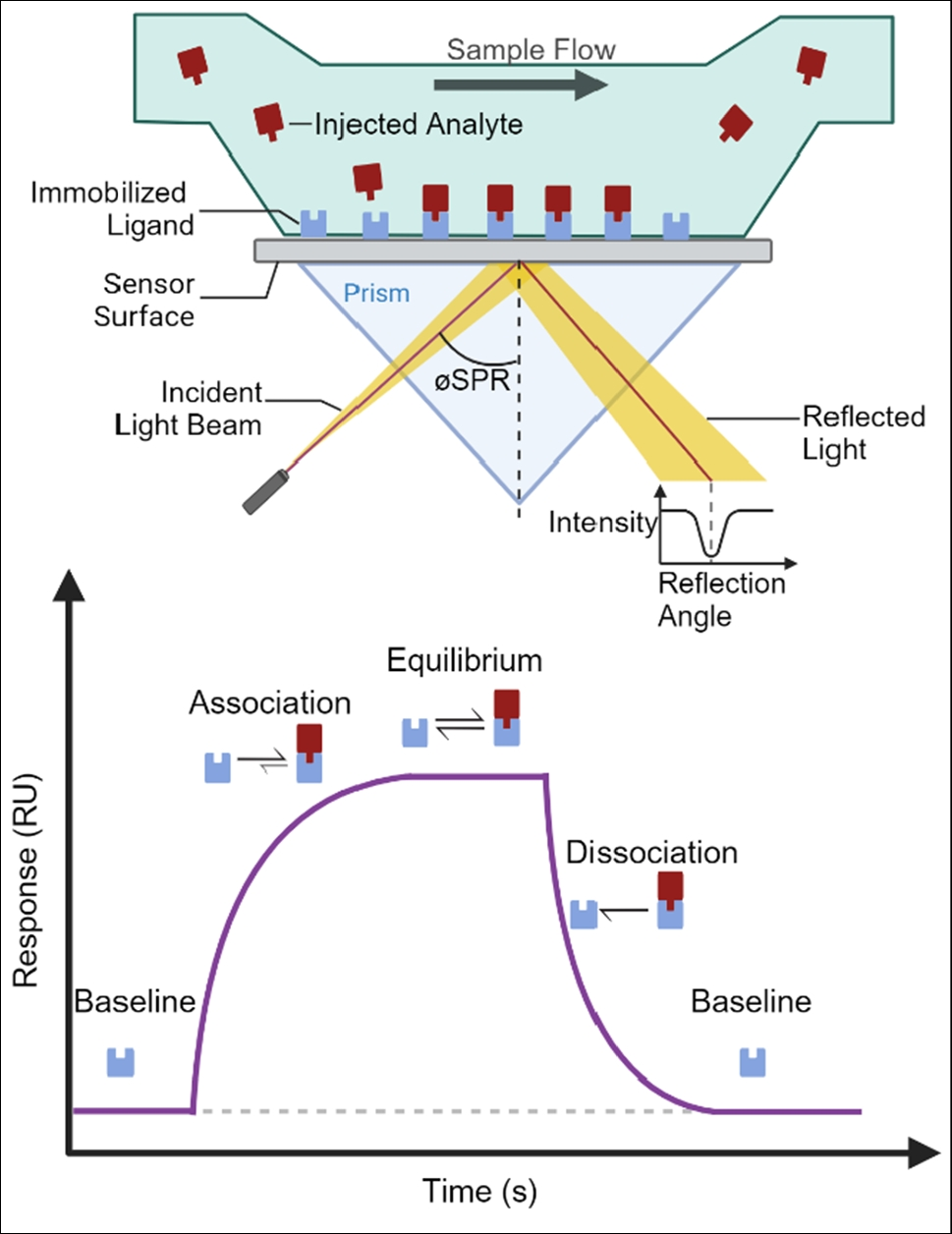
Table 2 Effects of Radopholus similis on Citrus limon growth 30 days after inoculation with 200 nematodes and the population density of the nematode recovered from rootTropism and invasion of 10 R. similis populations to the root system of rough lemonThe tropism test of 10 R. similis populations to the rough lemon root system showed that the nematodes of all populations were randomly and uniformly dispersed in the gel when the root segment was placed in the well, and the nematodes started to migrate and aggregate to the root segment after 2 h and 4 h. The nematodes of the SWK, GJ, FZ, GZ, DBSR, and YJ populations were observed to invade the root from the wound position after 6 h, whereas the nematodes of the XIN, ML, HN6, and HL populations were still around the root and did not invade it (Fig. 2A). The root tissues treated for 8 h were dyed with modified sodium chlorate-acid fuchsin. Under the stereomicroscope, many nematodes of SWK, GJ, FZ, GZ, DBSR, and YJ populations were observed in the root segments, but only one or no nematodes of the XIN, ML, HN6, and HL populations were found in the root segments (Fig. 2B). The results showed that all the tested populations tended towards the rough lemon root; however, the SWK, GJ, FZ, GZ, DBSR, and YJ populations could invade the rough lemon root, whereas the XIN, ML, HN6, and HL populations did not invade the rough lemon root.
Fig. 2
Tropism of 10 populations of Radopholus similis to the Citrus limon root. A: Tropism of R. similis to the root of C. limon in Pluronic F-127 gel evenly mixed with 200 female nematodes 6 h after assay initiation; B: Fuchsin staining of C. limon roots 8 h after assay initiation by infection with 200 female nematodes; XIN, ML, SWK, HN6, HL, GJ, FZ, GZ, YJ, and DBSR: the C. limon roots were infected with populations of R. similis, which originated from Zingiber officinale Roscoe (XIN), Crataegus pinnatifida (ML), Musa AAA Giant Cavendish cv.Baxi (HN6), Maranta arundinacea (HL), Chrysalidocarpus lutescens (SWK), Citrus reticulata (GJ), Anthurium andraeanum ‘Pink Champion’ (FZ), Anthurium andraeanum Linden (GZ), Curcuma longa (YJ), and Anubias nana (DBSR), respectively; scale bar = 500 μm
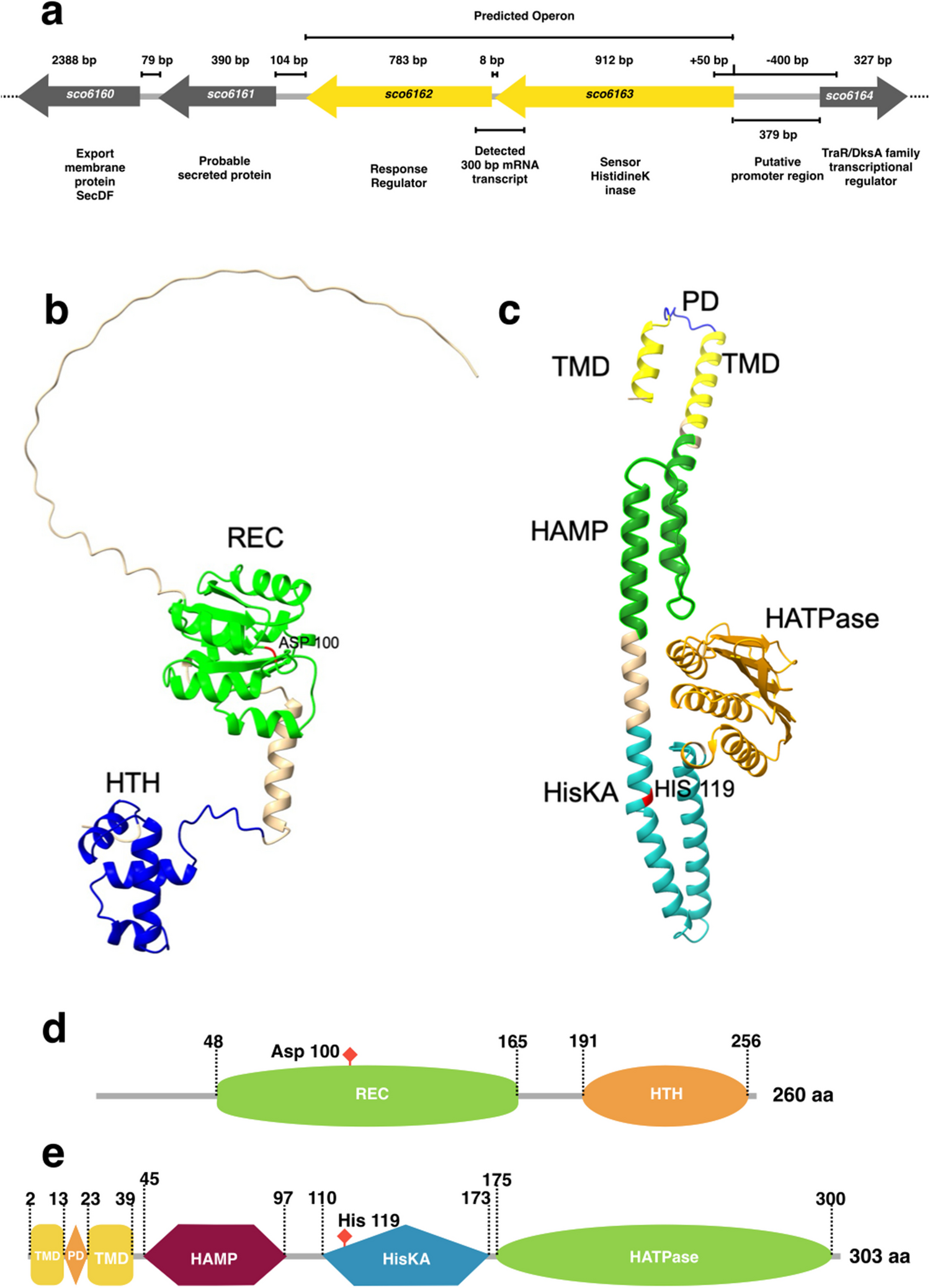
Cloning and analysis of genes of the pel family in R. similisFive transcripts of suspected pels were obtained via screening and blasting the transcriptome data of the GJ population of R. similis. The ORF and DNA genomic sequences of these genes were obtained using PCR amplification (Fig. 3).
Fig. 3
PCR amplification of five pectate lyase genes from Radopholus similis (Rs-pels). A: Amplification of the ORF sequences of Rs-pels; B: Amplification of the genomic DNA sequences of Rs-pels. M:DS2000 marker (GDSBio Co., Ltd., Guangzhou China); 1–5: products of Rs-pel-1, Rs-pel-2, Rs-pel-3, Rs-pel-4, Rs-pel-5 amplification respectively
After sequencing these fragments, a BLAST comparison confirmed that the deduced proteins of all five genes had conserved domains of the PEL family, and these five pels were named Rs-pel-1, Rs-pel-2, Rs-pel-3, Rs-pel-4, and Rs-pel-5. Their ORFs were 843, 819, 873, 771, and 810 bp long, and the number of encoded amino acids (aa) were 280, 272, 290, 256, and 269 aa, respectively (Table 3). ClustalX was used to compare the amino acid sequences of these five Rs-PELs with known Class III PEL sequences, and Rs-PEL-2, Rs-PEL-3, Rs-PEL-4, and Rs-PEL-5 were confirmed to have four conserved regions unique to the Class III PEL family (Fig. 4) (Shevchik et al. 1997), whereas Rs-PEL-1 did not have a conserved region unique to Class III PEL.
Table 3 Sequence analyses of pectate lyases of Radopholus similisFig. 4
Predicting the conserved regions of pectate lyases of the Class III PEL family from Radophulus similis. Regions I–IV indicate conserved regions characteristic of the Class III pectate lyase family (Shevchik et al.1997). Highly conserved charged residues are indicated with asterisks (*), RS_PEL2 to 5 indicate the amino acid sequences of Rs-PEL-2 to Rs-PEL-5 from R. similis, the species of the bacterium or the fungus, the gene, the amino acid size, and the accession number of the aligned sequences are: F_sol_PelA = Fusarium solani f. sp. pisi, PelA, 242 aa (M94692.1); F_sol_PelB = F. solani f. sp. pisi, PelB, 242 aa (U13051); F_sol_PelC = F. solani f. sp. pisi, PelC, 215 aa (U13049); F_sol_PelD = F. solani f. sp. pisi, PelD, 233 aa (U13050); E_car_PelB = Erwinia carotovora, PelB, 347 aa (X79232)
Sequence analysis of the five Rs-PELs predicted that none of them had a transmembrane domain. Among them, Rs-PEL-2, Rs-PEL-4, and Rs-PEL-5 contained signal peptides, whereas Rs-PEL-1 and Rs-PEL-3 did not have signal peptide sequences (Table 3).
Amino acid sequences of the five Rs-PELs were blasted against those of other plant-parasitic nematodes, bacteria, and fungi, and a phylogenetic tree was constructed using the maximum likelihood method in MEGA software (Fig. 5). The results showed that PELs from fungi and bacteria clustered into a large category, whereas those from plant-parasitic nematodes clustered independently into another large category. The Rs-PELs could be divided into three categories: first, Rs-PEL-3 and the PELs from four species of Meloidogyne were clustered into a branch; second, Rs-PEL-2 and Rs-PEL-5 were clustered with the PELs from two species of Meloidogyne and species of Heterodera and Globodera into a branch; third, Rs-PEL-1 and Rs-PEL-4 were clustered with the PELs from the nematodes of Heterodera, Globodera, and Aphelenchida into one branch.
Fig. 5
Maximum-likelihood phylogenetic trees of pectate lyases of Radopholus similis (Rs-PELs) and other organisms. Phylogenetic tree for proteins with conserved domains of pectate lyases from cyst nematodes, root-knot nematodes, Aphelenchus, Bursaphelenchus, bacteria, and fungi generated by MEGA6.0. The Rs-PELs amino acid sequences were marked in bold, and each sequence was followed by its accession number in GenBank
Expression of PEL genes in 10 populations of R. similisSingle nematode DNA from 10 populations was used as the template, and sequence amplification of four Rs-pels of the class III PEL family, i.e., Rs-pel-2, Rs-pel-3, Rs-pel-4, and Rs-pel-5, was performed. The results showed that Rs-pel-2, Rs-pel-4, and Rs-pel-5 could be amplified from the SWK, GJ, FZ, GZ, DBSR, and YJ populations that parasitised rough lemon, and the XIN, ML, HN6, and HL populations that did not parasitise rough lemon. However, the gene Rs-pel-3 could only be amplified from the six populations parasitizing rough lemon, showed stable expression in four developmental stages of these populations, and could not be amplified from the four non-parasitized rough lemon populations (Fig. 6A). Therefore, Rs-pel-3 could be used as a target gene to identify the banana and citrus pathotypes of R. similis and could be identified at all developmental stages in the citrus pathotype (Fig. 6B).
Fig. 6
DNA amplification of pectate lyase genes of Radopholus similis (Rs-pels) in 10 populations. A: DNA amplification of Rs-pels in 10 populations of R. similis; B: DNA amplification of Rs-pel-3 in different populations and developmental stages of R. similis; XIN, ML, SWK, HN6, HL, GJ, FZ, GZ, YJ, and DBSR: single nematode DNA from populations of R. similis, which originated from Zingiber officinale Roscoe (XIN), Crataegus pinnatifida (ML), Musa AAA Giant Cavendish cv.Baxi (HN6), Maranta arundinacea (HL), Chrysalidocarpus lutescens (SWK), Citrus reticulata (GJ), Anthurium andraeanum ‘Pink Champion’ (FZ), Anthurium andraeanum Linden (GZ), Curcuma longa (YJ), and Anubias nana (DBSR), respectively; M: DS2000 marker (GDSBio Co., Ltd., Guangzhou China); Fe: female; Ma: male; J2: second stage juvenile; Eg: egg
留言 (0)