
記住我
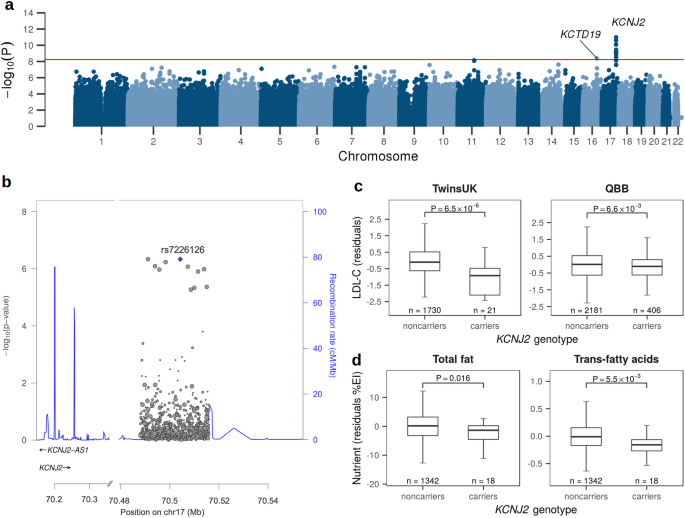

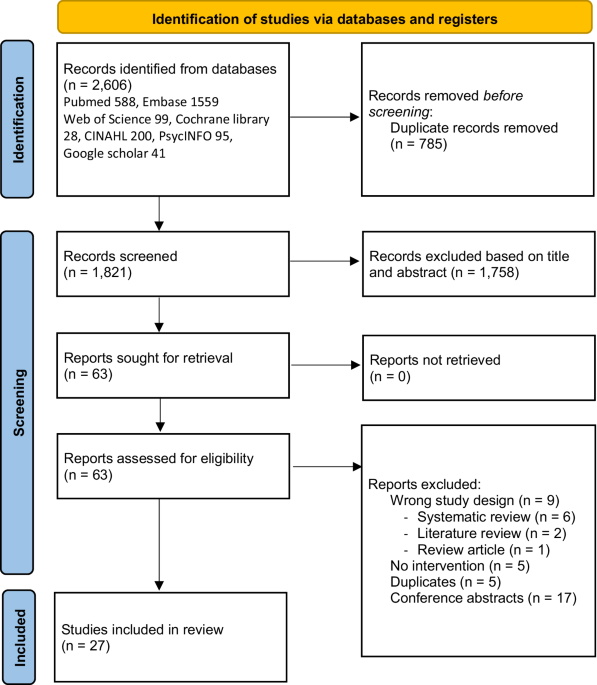
Since the initial publication of five families with pathogenic variants in BGN in 20171, thirteen additional families with BGN variants were identified (Fig. 1 and Table 1).
Fig. 1: Pedigrees of thirteen families with BGN variants identified in the current study.
Circle = female. Square = male. Diamond = unknown sex. Filled = patients with connective tissue features. Unfilled = unaffected or asymptomatic person. Strikethrough = deceased person. + = hemi-/heterozygous BGN variant carrier. (+) = heterozygous BGN variant carrier reported by proband. – = tested negative for BGN variant. Arrow = proband.
Table 1 Meester-Loeys syndrome cohort extension led to the identification of BGN variants in thirteen additional familiesFrameshiftIn families 1 and 2, likely pathogenic frameshift variants were identified (family 1: c.46delG, p.Ala16Profs*20, SCV004170959; family 2: c.59_60insAA, p.Gln21Serfs*16, SCV004170960). Skin fibroblasts were available from the proband of family 1 (1-III-3). Although we could not confirm the occurrence of nonsense-mediated mRNA decay (NMD), we did not observe any biglycan protein expression by western blot (Fig. 2). Another identical frameshift variant was identified in families 7 and 8 (c.441delinsAA, p.Asn148Lysfs*54, SCV004170964). This frameshift variant is present as two separate entries (rs782449715 and rs782199865) in GnomAD in three individuals22. Intriguingly, these three GnomAD individuals (one female and two males) are all part of the European Finnish population, and both probands of families 7 and 8 originate from (Northern) Finland. Western Blot on protein derived from skin fibroblasts of the male proband of family 7 (7-II-3) confirmed the predicted complete absence of the biglycan protein (Supplementary Fig. 1). Although a small fraction of mutant peaks was observed with cDNA analysis of the variant-harboring unaffected mother (7-I-1), no mutant or decreased concentration of biglycan protein was observed by Western Blot (Supplementary Fig. 1). No skin fibroblasts from family 8 were available for further testing of this BGN frameshift variant.
Fig. 2: Western Blot of BGN protein expression in skin fibroblasts of probands from families 1, 6, 9, 10, 17, and 18 as well as matched controls.
Intracellular proteins were isolated from skin fibroblast samples and the biglycan (BGN) protein content was visualized. Cyclophilin B (CypB) was used as a loading control. Family 17 and 18 were reported as family 4 and 5, respectively, by Meester et al.1. Controls 1–4 were samples of age-matched male controls. The Western Blot was derived from one experiment and all lanes were processed in parallel.
NonsenseIn the probands of families 3 to 5, likely pathogenic nonsense variants were identified (families 3 and 4: c.75 G > A, p.Trp25*, SCV004170961; family 5: c.223 C > T, p.Gln75*, SCV004170962), which are predicted to lead to NMD and biglycan protein absence. No skin fibroblasts of the probands or family members were available for further testing of this hypothesis.
MissenseIn family 9, a missense variant was identified (c.565 G > A, p.Glu189Lys, SCV004170965). Since this variant affects the last nucleotide of exon 4, and thus a canonical splice site, cDNA analysis of RNA derived from skin fibroblasts of the male proband (9-II-3) was performed to determine the effect of this variant. We observed an alternatively spliced transcript that lacks the last 46 nucleotides of exon 4 due to the use of an exonic, cryptic splice site, resulting in a frameshift effect (p.Val174Argfs*20) and that partly undergoes NMD. Furthermore, the wildtype spliced transcript (containing the missense variant) was also detected on the cDNA level. Nonetheless, no biglycan protein expression was observed by western blot (Fig. 2), confirming that this variant causes a complete loss of the biglycan protein.
Splice siteIn families 6 and 10 to 13, splice site variants were identified. The variant of family 6 affects the first nucleotide of intron 3 (c.351+1 G > A, SCV004170963). cDNA analysis of RNA derived from skin fibroblasts of the male proband (6-III-1) showed the use of a cryptic splice site in intron 3, leading to an in-frame insertion of 51 nucleotides and, consequently, an addition of 17 amino acids (p.Tyr117_Ala118insIleArgSerTrpGluGluProAlaGlyLeuGlnGlnArgAlaGlyValArg). The presence of this aberrant (longer) protein was also observed by Western Blot (Fig. 2), indicating that this mutant biglycan protein is not (completely) degraded. No normal biglycan protein was observed. In families 10 and 11, a splice site variant affecting the same nucleotide, but resulting in a different substitution, was identified (family 10: c.677-2 A > G, SCV004170966; family 11: c.677-2 A > T, SCV004170967). Skin fibroblasts of the male probands from these two families (10-II-2 and 11-II-1) were available for cDNA and Western Blot analysis. The BGN variants identified in families 10 and 11 were shown to result in the activation of a cryptic splice site in exon 6, leading to the loss of the first 33 nucleotides of that exon, and, consequently, the in-frame loss of 11 amino acids (p.Asp225_Leu236del). This aberrant shorter biglycan protein was confirmed by Western Blot (Fig. 2 and Supplementary Fig. 1). No normal biglycan protein was observed. The variants identified in families 12 and 13 affect the canonical donor splice site of exon 6 and the acceptor splice site of exon 8, respectively (family 12: c.770+1 G > A, SCV004170968; family 13: c.910-1 G > A, SCV004170969). Although no skin fibroblasts were available for further testing, splice prediction algorithms predict distinct effects on splicing. For family 12, the donor splice site is predicted to be lost, potentially leading to the skipping of exon 6, which would result in a frameshift. For family 13, a novel splice site is predicted to be located one nucleotide further downstream than the canonical splice site, also leading to a frameshift. Since exon 8 is the last exon of Biglycan, no NMD is predicted to occur. Yet, the N-terminal protein structure would be altered because of the variant.
Further investigation of previously identified BGN variantsThe following two families (17 and 18) were already reported by Meester et al.1, but in this study, we performed additional assays to determine the effect of the different reported BGN variants on protein level by Western Blot. The male proband of family 17 (family 4 in Meester et al.1) carries a 28 kb deletion of the coding part of biglycan (ChrX(GRCh38):g.153502980_153530518del; VCV000265797.1). Western Blot experiments confirmed the absence of the biglycan protein in this patient (Fig. 2). The splice site variant in family 18 (family 5 in Meester et al.1; c.238 G > A, p.Gly80Ser; VCV000265798.2) was described to result in four different splice products, of which two underwent NMD. The other two, less abundant, splice products did not lead to NMD. However, no biglycan protein expression was observed by western blot (Fig. 2), indicating a complete loss of the biglycan protein in this male patient (18-II-1).
BGN 5’-UTR hijacking by ATP2B3The 28 kb deletion in the proband of family 17 (17-II-1; family 4 in Meester et al.1) abolishes the coding part of BGN (exon 2-8). In addition, the deletion affects a portion of DNA downstream of BGN, containing the start of the 5’ untranslated region (5’-UTR) of several ATP2B3 isoforms. RNA sequencing on the skin fibroblasts of this patient was performed to investigate the effect of this deletion in more detail. As expected, no expression of the coding exons of BGN was observed (Fig. 3, red box). However, the 5’-UTR of BGN remained expressed (Fig. 3, green box). Interestingly, due to the partial loss of its own 5’-UTR (Fig. 3, orange box), ATP2B3 now uses the 5’-UTR (exon 1; Fig. 3, green box) of BGN to drive its expression (Fig. 3, blue box). ATP2B3 is not expressed in healthy control skin fibroblasts, but due to hijacking of the 5’-UTR of BGN, it is now expressed in this cell type as a consequence of the 28 kb deletion in this patient.
Fig. 3: mRNA sequencing of skin fibroblasts of the proband from family 17 (17-II-1) shows BGN 5’-UTR hijacking by ATP2B3.
Family 17 was reported as family 4 by Meester et al. (2017)1. The control sample was age- and sex-matched.
Cohort characteristicsCombining the previously published cohort1 with the additional thirteen families, our cohort now includes 18 probands (16 males and 2 females) with an average age at presentation of 33 years, ranging from 0 to 70 years. Thirteen male probands presented with aortic (n = 10) and/or widespread arterial (n = 8) aneurysms/dissections, a male newborn (4-III-3, 0.5 years) presented with marked hydrocephaly, craniofacial features, pectus excavatum and syndactyly, a tall male (6-III-1, 36 years) with significant camptodactyly, spine deformities, flat feet, and joint contractures, and another male (7-II-3, 13 years) presented with various syndromic findings but without cardiovascular features (yet). Our cohort comprises two female probands: one (13-III-2, 59 years) presented with an aortic aneurysm, whereas the other (5-II-2, 44 years) was detected as part of a comprehensive prenatal testing study. Segregation analyses identified 36 additional BGN variant-harboring family members (9 males and 27 females). The clinical phenotype of female BGN variant carriers ranged from no phenotype to aortic aneurysm with typical MRLS connective tissue features. A summary of the clinical features of all BGN variant-harboring individuals can be found in Table 2. A detailed overview of the clinical features of the individuals from families 1 to 13 can be found in Supplementary Table 1 and the Supplementary Notes containing the family case reports, and for families 14 to 18 in Meester et al.1.
Table 2 Clinical features of all BGN variant-harboring individuals (n = 54) from extended Meester-Loeys syndrome cohortCardiovascular featuresImportant cardiovascular features, all with a predominance in males, include aortic root aneurysm (35%; M/F: 56%/17%), ascending aortic aneurysm (14%; M/F: 26%/4%), and aortic dissection (8%; M/F: 14%/4%). Strikingly, 25% of the BGN variant-harboring individuals had arterial aneurysms (M/F: 44%/10%; Supplementary Fig. 2a, b) and 13% arterial dissections (M/F: 31%/0%). Both features occurred more frequently than originally described1, and showed a clear predominance in males.
Skeletal featuresA wide range of skeletal features was observed in our MRLS cohort. Joint hypermobility (47%; M/F: 57%/38%), reduced bone density (46%; M/F: 57%/33%), and flat feet (36%; M/F: 47%/25%) were observed most often. Other recurrent skeletal features include pectus deformity, spine deformity, spatulous fingers, joint dislocation, and joint contractures. Both, short and tall statures were noted. Similarly, both, arachnodactyly and brachydactyly (Supplementary Fig. 2d) were observed.
Craniofacial featuresThe most common craniofacial features in our MRLS cohort include malar hypoplasia (26%; M/F: 39%/15%), downslanting palpebral fissures (24%; M/F: 35%/14%), and hypertelorism (20%; M/F: 30%/13%; Supplementary Fig. 2c). Other recurrent, but less frequent, craniofacial features were dolichocephaly, proptosis and gingival hypertrophy. Strikingly, broad or bifid uvula, highly arched palate, and frontal bossing were only noted in males.
Other featuresMyopia was the only recurrent ocular feature, noted in 30% (M/F: 33%/28%). Mild learning problems (11%; M/F: 25%/0%) and ventriculomegaly (27%; M/F: 36%/0%) only occurred in males, while relative macrocephaly or large head circumference was noted for both sexes (35%; M/F: 50%/13%). Dermatomyositis was noted for one female proband. Regarding cutaneous manifestations, striae (18%; M/F: 11%/25%) and easy bruising (11%; M/F: 7%/15%) were observed more frequently in females than in males. Other cutaneous findings included hypertrichosis, delayed wound healing, and umbilical hernia.
留言 (0)