
記住我
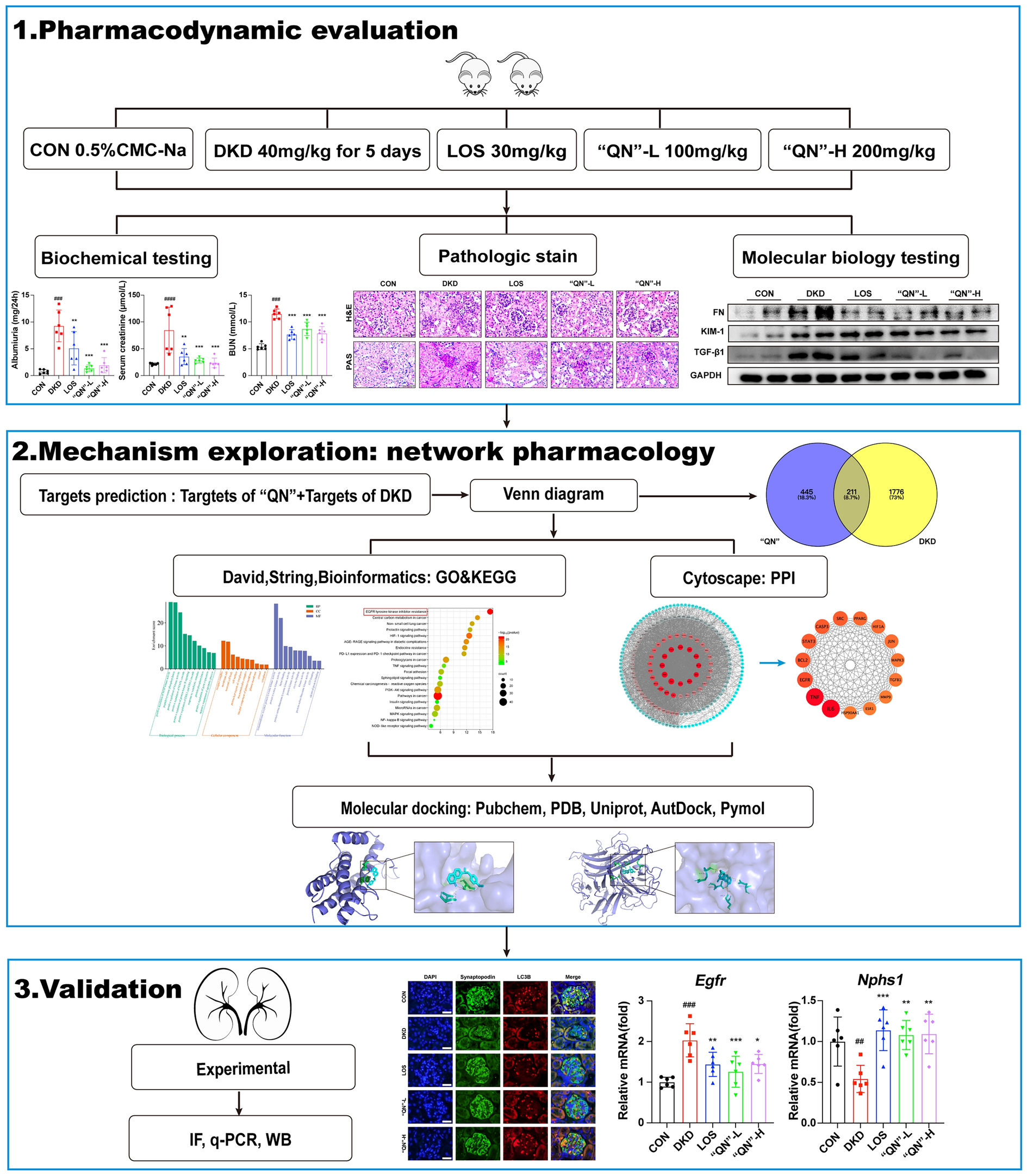



To determine a noncytotoxic concentration suitable for the anti-HBV study, we assessed the cytotoxic effects of LWWL on HepG2.2.15 cells. Our findings revealed that when the concentration of LWWL was below 2 mg/ml, the rate of cell death remained below 5% (Fig. 1a). This indicates that LWWL at a concentration of 2 mg/ml or lower exhibited minimal cytotoxicity towards the cells. Consequently, a maximum concentration of 2 mg/ml of LWWL was employed in this study. After being treated with LWWL for 24 h, the levels of HBsAg and HBeAg in the culture media were then assessed using ELISA. The results demonstrated that LWWL effectively decreased HepG2.2.15 cell production of HBsAg, HBeAg, and HBV DNA. Furthermore, the inhibition exhibited a dose-dependent relationship (Fig. 1b–d). To sum up, these results support LWWL’s anti-HBV activity in vitro.
Fig. 1
LWWL inhibited the replication of HBV in vitro. HepG2.2.15 cells were treated with the indicated concentrations of LWWL for 24 h. Cell death was measured by CellTiter-Glo luminescent assay (A). HepG2.2.15 cells were treated with the indicated concentrations of LWWL for 24 h. Then the supernatant was collected to detect the levels of HBeAg, HBsAg, HBV DNA (B–D). The data are presented as mean ± SEM from triplicates, *P < 0.05, **P < 0.01, ***P < 0.001, NS: not significant (one-way ANOVA with Dunnett’s post-hoc test). HepG2.2.15 cells were subjected to treatment with varying concentrations of LWWL for a duration of 24 h. Cell death was assessed using the CellTiter-Glo luminescent assay (A). HepG2.2.15 cells were exposed to the respective concentrations of LWWL for 24 h, and the supernatant was collected for analysis of HBeAg, HBsAg, and HBV DNA levels (B–D). The data presented are mean values ± standard error of the mean (SEM) from triplicates, *P < 0.05, **P < 0.01, ***P < 0.001, and NS denotes not significant (one-way ANOVA with Dunnett’s post-hoc test)
Building on this foundation, we have further confirmed whether the combination of LWWL and ETV is better than the single use, we tested a series of viral marker on hepatitis B virus, such as HBsAg, HBeAg and so on. The results showed a significant decrease in the levels of both HBeAg and HBV DNA in the serum of mice treated with LWWL, ETV or their combination (Fig. 2a–f). Furthermore, the level of HBsAg in the serum of mice treated with LWWL also decreased (Fig. 2g, h). Compared to the monotherapy group, the combination group showed a better decrease in the levels of HBeAg and HBV DNA in the serum. It was indicated that the synergistic inhibitory effect of the combination of LWWL and ETV on hepatitis B virus was significantly superior to that of treating with LWWL or ETV alone. Consistent with the immunohistochemical assessment, the combination treatment was more effective in reducing HBcAg in tissues compared to individual treatment as expected (Fig. 2i).
Fig. 2
The combination of LWWL and ETV reduces hepatitis B virus replication in vivo. C57BL/6J mice were hydrodynamic injection of pAAV/HBV1.2 plasmids at a dose of 20 μg through the tail vein within a duration of 6–8 s, controls were injected with saline equivalent to 10% of the mouse’s body weight. After 3 days, blood was taken from the orbit to detect the levels of HBsAg, HBeAg and HBVDNA in the serum. Upon observing an increase in the expression of HBsAg, HBeAg, and HBV DNA in the mice, the mice injected with plasmids were divided into four groups (n = 6). These groups received treatment with saline, LWWL, ETV, and a combination of LWWL and ETV, respectively, at concentrations below the minimal liver-cytotoxic level for a period of 20 days. The control group was administered saline. The levels of serum HBeAg (A–C), HBsAg (D–F), and HBV DNA (G, H) were measured at specified time points. At the end of the 20-day period, the mice were sacrificed, and the expression of HBcAg in the liver was assessed using immunohistochemistry (D). The scale bars represent 100 μm (top row) and 20 μm (bottom row). Data are shown as mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, NS: not significant (one-way ANOVA with Dunnett’s post-hoc test)
The results demonstrated that compared to the single use, the combination of LWWL and ETV had a greater antiviral effect in vivo.
Network pharmacology-based analysisIn order to investigate the main components of LWWL for the treatment of hepatitis B, we conducted a network pharmacology analysis. Within the TCMSP database, a total of 36 active compounds of LWWL were identified based on the criteria of OB ≥ 30 and DL ≥ 0.18. Additionally, the main compounds of Qu Maicai and Lingzhi Baozifen were supplemented based on relevant literature. Consequently, we obtained a total of 50 active ingredients of LWWL (detailed compound information is provided in Additional file 1: Tables S1 and S2). The active ingredients of LWWL were further matched to potential targets, resulting in the inclusion of a total of 250 targets.
Through the utilization of the following 5 databases, namely DisGeNET, GeneCards, OMIM, TTD, and Drugbank, we identified 1449, 474, 593, 20, and 39 confirmed or potential HBV targets, respectively. To ensure consistency, all targets were standardized using Uniprot. Duplicate targets were subsequently removed, resulting in a final set of 2000 targets.
The intersection of active ingredients and disease targets was visualized using a Venn diagram (Fig. 3a), which revealed 109 common targets. As a result, we discovered that the 37 active compounds were associated with 109 HBV-related targets. To visualize the LWWL active ingredient-target-disease network, we utilized Cytoscape 3.9.1. The network consisted of 152 nodes and 350 edges (Fig. 3b). Analysis of the network demonstrated a centrality value of 0.55 and a heterogeneity value of 1.89, illustrating the synergistic therapeutic effect of LWWL through the interaction of multiple ingredients with multiple targets. Detailed information on the top 10 compounds is provided in Table 2. Also, 109 intersection targets in the figure were input into STRING11.0, and got 102 targets and made them a PPI network diagram of LWWL for HBV by Cytoscape (Fig. 3c).
Fig. 3
Network pharmacological analysis of LWWL on hepatitis B virus treatment. A Venn diagram of LWWL and HBV therapeutic targets. B The LWWL-Ingredients-Target-HBV network. C Protein–protein interaction network of common targets of LWWL and HBV. D GO enrichment analysis. E KEGG enrichment analysis
Table 2 Compounds in the ingredient-target-disease network of Liuwei Wuling Tablets (top 10)Metascape was utilized to conduct GO analysis of targets in PPI network. It shows that the numbers of cellular components (CC), molecular functions (MF), and biological processes (BP) are 65, 125, 15,900 respectively (P < 0.05, the bar graph of the top 10 is shown in Fig. 3d). Following KEGG Pathway analysis (P < 0.05), we identified a total of 180 signaling pathways. Top 6 signaling pathways are: Pathways in cancer, AGE-RAGE signaling pathway in diabetic complications, Lipid and atherosclerosis, Hepatitis B, PI3K-Akt signaling pathway, MAPK signaling pathway, (the bubble chart of the top 20 is shown in Fig. 3e).
Lut inhibits HBV replication in HepG2.2.15 cells via ERK-mediated downregulation of HNF4aIn order to investigate the antiviral effects of compounds in LWWL, the top five drugs were chosen based on network pharmacology degrees, and their effects on the concentrations of HBeAg in the HepG2.2.15 cells’ supernatant were examined by using ELISA. The data demonstrated a significant reduction in the level of HBeAg by luteolin (Fig. 4a). It suggests that luteolin (Lut) may have antiviral effect in vitro.
Fig. 4
Lut inhibits the expression of HNF4α and HBV replication through activating MAPK/ERK pathway. HepG2.2.15 cells were treated with indicated compounds for 24 h. Then the supernatant was collected to detect the levels of HBeAg (A). HepG2.2.15 cells were treated with the indicated concentrations of Lut for 48 h. Cell death was measured by CellTiter-Glo luminescent assay (B). HepG2.2.15 cells were treated with the indicated concentrations of Lut for 24 or 48 h. then the supernatant was collected for the determination of HBeAg, HBsAg, HBV DNA (C–E). HepG2.2.15 cells were treated with Lut (5, 10, 20 and 40 μM) for 6 h, the mRNA was acquired for the measurement of HNF4α (F) by qRT-PCR. HepG2.2.15 cells were incubated with Lut (5, 10, 20 and 40 μM) for 24 h, and the blot intensity of p-ERK1/2 (G) was analyzed through Western blot. The data are presented as mean ± SEM from triplicates, *P < 0.05, **P < 0.01, ***P < 0.001, NS: not significant (one-way ANOVA with Dunnett’s post-hoc test)
For further study of evaluating the antiviral ability of Lut in vitro, we initially assessed the cytotoxicity of Lut in HepG2.2.15 cells using the CellTiter-Glo luminescent assay. Following a 48-h exposure of HepG2.2.15 cells to Lut, the cell viability was measured, and the results indicated that concentrations of Lut below 40 µM resulted in a cell death rate of less than 5%, suggesting that Lut at a concentration of 40 µM or lower exhibited minimal cytotoxicity (Fig. 4b). Therefore, Lut with a maximum concentration of 40 µM was used. Subsequently, the impact of Lut on HBV replication in HepG2.2.15 cells was examined. After treating Lut for 24 and 48 h, the levels of HBsAg and HBeAg in the culture media were assessed. The results revealed that Lut inhibited the release of HBsAg and HBeAg from HepG2.2.15 cells (Fig. 4c, d). The inhibitory effects of Lut on HBsAg and HBeAg were dose-dependent and found to increase with the concentration of luteolin used, indicating a clear dose-dependent manner, also enhanced with time in HepG2.2.15 cells. Also, the level of HBV DNA was evaluated, it showed the inhibition of HBV DNA replication was significant within the concentration range from 10 to 40 μM of luteolin (Fig. 4e).
Hepatocyte nuclear factor 4α (HNF4α) is a member of the HNF family, plays a key role in HBV transcription, particularly in the generation of pgRNA [35, 36]. The activation of the MAPK signaling pathway has the potential to disturb the communication between enhancers and promoters of HNF4α, resulting in the down-regulation of HNF4α expression [37]. It has been reported before that Lut could activate MAPK/ERK pathway to downregulate the expression of HNF4α [30], we have confirmed Lut significantly increase the phosphorylation of ERK (p-ERK) exhibited a dose-dependent manner (Fig. 4f, g), and inhibited the expression of HNF 4 α in a dose-dependent manner.
Overall, the findings revealed that Lut significantly inhibited HBV replication via ERK-mediated downregulation of HNF4α.
SC promotes the activation of cGAS-STING pathway and interferon production in THP-1 cells to suppress HBV replication in HepG2.2.15 cellsIn addition to direct antiviral drugs, there is a growing interest in the role of immunity which consist of adaptive and innate immunity in the treatment of HBV. cGAS-STING pathway, one of the important antiviral pathways [38, 39], has been reported to play an essential role in inhibiting HBV. SC, one of the main lignan of Schisandra chinensis (sovereign drug of LWWL), has been reported to facilitate the activation of cGAS-STING pathway in macrophages [32].
We proposed that SC augmented the activation of the cGAS-STING pathway in macrophages to induce interferon secretion, thereby suppressing the replication of HBV DNA in hepatocytes and achieving an antiviral effect.
To verify this hypothesis, we established a co-culture system of THP-1 and HepG2.2.15 cells. We put transwell chamber which seeded with HepG2.2.2.15 cells on the top of 6-well transwell dish containing THP-1 cells pretreated with SC or vehicle followed by ISD (Interferon Stimulatory DNA) transfection or not. After 24 h we collected the supernatant to measure the levels of HBsAg and HBeAg. The datas showed that in the context of ISD transfection, the HBeAg and HBsAg’s level of the co-culture group were obviously reduced, and there was a better reduction in the group given with SC (Fig. 5b, c). Also, we evaluated whether SC has a direct antiviral effect on HepG2.2.15 cells. The results indicated that treating HepG2.2.15 cells with SC did not lead to a decrease in the levels of HBsAg and HBeAg.
Fig. 5
SC has an antiviral effect in vitro by increasing the production of IFN via ISD-induced cGAS-STING pathway activation. PMA-primed THP-1 cells which were seeded in the 6-well plate were treated with SC (20 µM) for 2 h, then stimulated with ISD for 2 h. Put Transwell chamber which seeded with HepG2.2.2.15 cells on the top of 6-well transwell dish which seeded with THP-1 cells (A). After 24 h we collected the supernatant to measure the levels of HBeAg and HBsAg (B, C). PMA-primed THP-1 cells were incubated with SC (5, 10, and 20 μM) for 2 h, then stimulated with ISD for 2 h and the blot intensity of p-IRF3, STING, IRF3 (D) was analyzed through Western blot. PMA-primed THP-1 cells were treated with DMSO or SC for 2 h, then stimulated with ISD for 3 h, induction of IFN β mRNA was measured by qRT-PCR (E). PMA-primed THP-1 cells were treated with DMSO or SC for 2 h, then stimulated with ISD for 6 h, secreted IFN β were detected by ELISA (E). Data are shown as mean ± SEM from triplicates, **P < 0.01, ***P < 0.001, NS: not significant (one-way ANOVA with Dunnett’s post-hoc test or unpaired Student’s t-test)
Next, we confirmed the effect of SC on the cGAS-STING signaling pathway, the results showed that the phosphorylation of IRF3 (p-IRF3) exhibited a dose-dependent increase (Fig. 5c). And the mRNA expression of interferon and the expression of IFN-β in the supernatant of THP-1 cells were evaluated. As anticipated, the expression of mRNA and interferon in the supernatant increased with the rise in concentration (Fig. 5d, e).
Above all, the results could suggest that SC can raise cGAS-STING pathway’s activation, resulting in the secretion of IFN-β and thereby decreasing the levels of HBsAg and HBeAg in the supernatant.
A combination of SC and Lut significantly inhibits HBV DNA replication in vivoPrevious experiments have indicated that luteolin can suppress HBV replication by downregulating HNF4α through the MAPK/ERK pathway, while SC can directly promote innate immunity and enhance the activation of the cGAS-STING antiviral pathway to inhibit viral replication. Subsequently, we proceeded to assess whether the synergistic effect of SC and Lut demonstrated a superior anti-HBV activity when compared to their individual treatments in the HBV-infected mouse model. The PAAV/HBV1.2 plasmids was used to induce hepatitis B (Fig. 6a). After the injection of plasmid into the tail vein of mice, the level of HBsAg, HBeAg in the serum increased (Fig. 6d, g), indicating that the mice had been infected with hepatitis B virus, this was further confirmed by HBV DNA (Fig. 6b).
Fig. 6
The combination of SC and Lut reduces hepatitis B virus replication in vivo. C57BL/6J mice were hydrodynamic injection with 20 μg of pAAV/HBV1.2 plasmids through the tail vein within 6 − 8 s, controls were injected with saline equivalent to 10% of the mouse’s body weight. After 3 days, blood was taken from the orbit to detect the level of HBsAg, HBeAg and HBVDNA in the serum. When the expression of HBsAg, HBeAg, and HBV DNA in mice increases, the mice injected with plasmids were divided into 4 groups (n = 6), which were treated with saline, Lut, SC, and their combination under the minimal liver-cytotoxic concentration within 20 days, respectively (A). The control group was given saline. Serum HBeAg (D–F), HBsAg (G–I) and HBV DNA (B, C) levels were measured for indicated times. Mice were sacrificed at the 20 day, the expression of HBcAg in liver were detected by immunohistochemistry (J), scale bars: 250 μm (top row); 50 μm (bottom row). Data are shown as mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, NS: not significant (one-way ANOVA with Dunnett’s post-hoc test)
After that, the mice were randomly divided into 4 groups, except for the control group, and received gavages with saline (model group), SC (20 mg/kg), Lut (20 mg/kg), a combination of SC (20 mg/kg) and Lut (20 mg/kg) within 20 days. The data showed SC, Lut, and their combined treatment groups showed good effects in preventing hepatic B, and Inhibiting HBV replication, which the decrease in the levels of HBsAg and HBeAg in the serum can prove. Also, the combined group also showed better inhibition of HBsAg and HBeAg in the serum to the single drug group (Fig. 6e, f, h, i). Moreover, the coordinate repression of SC and Lut on reduction of HBV DNA in the serum was significantly superior than SC or Lut alone (Fig. 6c). HBcAg is an endonuclear protein of the hepatitis B virus, which plays an important role in the replication and assembly of the hepatitis B virus, and its test can be used for the diagnosis of hepatitis B, disease monitoring and prognosis assessment. Consistent with the results of liver histopathological evaluation, the combination therapy of SC and Lut also reduced the content of HBcAg in liver tissue more effectively than SC and Lut alone (Fig. 6j).
Thus, the combination of SC with Lut offered synergistic effects against the chronic hepatitis B.
留言 (0)