IRDs are a group of ophthalmic hereditary diseases with high genetic and clinical heterogeneity, and genetic testing has greatly assisted the clinical diagnosis of IRDs. A series of studies have demonstrated the contribution of SVs and intronic variants to the genetic diagnosis of IRDs17,20,21. In our current study, we identified 27 pathogenic SVs and intronic variants across 14 different IRD genes in 34 previously unresolved IRD cases through WGS. It is worth noting that all variants detected in these patients have not been previously reported, thus our study represents the first report of these specific variants, underscores the significance of SVs and intronic variants in IRDs.
In 271 IRD patients, the disease-associated SVs were identified in 11% of cases, in concordance with previous reports of the pathogenic proportion (5% to 15%) of SVs in the molecular diagnosis of IRDs15. It is worth noting that all cases included in this study had been previously screened by panel-based sequencing and showed negative results. Although panel sequencing also included CNV analysis, no large deletions or duplications suspected to be pathogenic were found in these cases. This underscores the potential of WGS as an optimal sequencing method for detecting SVs, offering a higher rate of genetic diagnosis. Notably, we observed that 4% of cases exhibited a compound heterozygous pathogenicity pattern involving both SVs and SNVs in recessive genes, highlighting a previously underexplored form of pathogenicity arising from the combined effects of SVs and SNVs. Such pathogenic patterns are often overlooked as SNV/indel and SV screening are typically conducted separately. Furthermore, in this study, intronic variants contributed an additional 2% to the overall pathogenicity, further affirming the diagnostic significance of deleterious variants located in non-coding regions, particularly those affecting mRNA splicing. Notably, all five of the intronic variants we identified had not been previously reported, suggesting that the contribution of deep-intronic variants to the molecular diagnosis of IRDs may have been underestimated in previous studies.
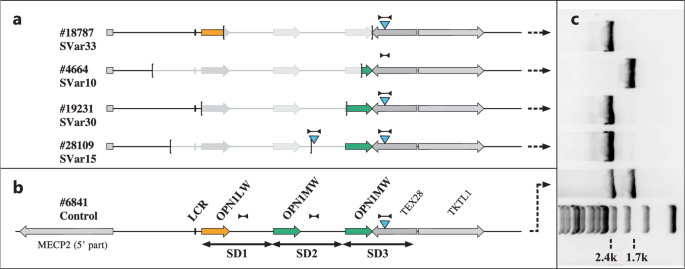
In total, 22 disease-associated SVs, 3 deep-intronic variants, and 2 non-canonical splice-site variants of IRD genes were found. Except for one inversion of ABHD12, SVs in the remaining 10 IRD genes were deletions (including homozygous, heterozygous, and hemizygous deletions), ranging from single exon to complete gene deletions. All detected SVs were defined as novel pathogenic variants, further extending the mutational spectrum of IRD genes. In this study, we confirmed that approximately three-quarters of SVs were clustered in RP-related genes, of which the most frequently altered genes by SVs were EYS (n = 5) and PRPF31 (n = 4). Previous studies have highlighted PRPF31, EYS, and USH2A as the most prevalent pathogenic genes harboring SVs in IRDs10. A homozygous deletion (chr4:633534–637421) in PDE6B was reported for the first time in this study, which was concurrently observed in three RP patients from two unrelated families (FM13 and FM105), corroborating the pathogenic significance of this deletion. Moreover, 5 novel intronic variants, including deep-intronic variants and non-canonical splice-site variants, were detected from 4 IRD genes in 4 families and one sporadic case. These intronic variants identified were validated to cause aberrant splicing by minigene assays.
None of the detected pathogenic intronic variants and SVs were observed or had extremely low AFs in the general population. The AFs of SVs varied widely among different population groups, with a very limited sharing of SVs between European and East Asian populations (Supplementary Fig. 1). We therefore used diverse background population groups to confirm true rare SVs, as a prerequisite for establishing pathogenicity. Using an inadequate population background can lead to misinterpretation of rare variants, especially when analyzing small, sporadic cohorts. We therefore recommend the SV reference set to include at least one population group matching the patient cohort. Notably, most current SV annotation methods do not consider tissue-specific transcript information. Consequently, benign SVs that do not affect specific transcripts in disease-relevant tissues may be frequently mis-labeled as pathogenic. As shown in our study, pathogenic and benign SVs exhibited rather different transcript disruption ratios. Implementing a threshold of 20% for transcript disruption ratio significantly reduced false positives in the identification of pathogenic SV identification, particularly in sporadic cases.
We also conducted genotype-phenotype correlations among IRD patients. The average age of the probands was 35 years (range 8–58 years old) and the average age of onset was 15.0 years (range 2–50 years old). Variants in the USH2A gene result in either RP (OMIM 613809) or Usher syndrome (OMIM 276901)22. In our study, the ocular phenotypes of patients harboring USH2A variants were consistent with clinical manifestations of RP, which were characterized by progressive night blindness and reduced visual field. The fundus showed waxy optic disc, retinal osteocytes-like pigmentation, retinal vascular stenosis, accompanied by retinal atrophy and thinning. However, these patients did not display symptoms of diseases beyond ocular involvement and were ultimately diagnosed with RP rather than Usher syndrome. By contrast, patients carrying MYO7A variants exhibited both RP and hearing impairment, aligning with the diagnosis of Usher syndrome. Notably, the proband with ABHD12 variants in FM2 had syndromic features, presenting with both characteristics of RP and deafness. We further checked the clinical phenotype of the remaining patients carrying variants in RP causative genes, and their symptoms and fundus manifestations were consistent with the clinical diagnosis of RP. The representative RP photographs from the proband in FM110 were shown in Supplementary Fig. 5. We detected variants in genes associated with CRD (DRAM2 and RIMS1) in two families, FM77 and FM134. The probands presented with decreased visual acuity and abnormal color vision, with electroretinography (ERG) indicating more severe impairment of cone photoreceptor function than rod photoreceptor. Additionally, fundus examinations revealed macular atrophy. For example, the color fundus photograph of proband from FM134 displayed waxy optic disc discoloration, macular atrophy, and retinal vessel narrowing. Fundus autofluorescence (FAF) showed hypo-autofluorescence in the macular area surrounded by a hyperfluorescent ring. Spectral domain optical coherence tomography (SD-OCT) examinations revealed thinning of the macular fovea thickness, particularly in the neuroepithelial layer (Supplementary Fig. 6). According to the genetic test results of the probands, combined with the clinical phenotype and medical history, it was consistent with the diagnosis of CRD.
The clinical manifestations of IRDs are diverse, and variants in the same causative gene can lead to different clinical phenotypes9, posing challenges in the genetic diagnosis of IRD patients. Variants in PROM1 are responsible for autosomal recessive or autosomal dominant IRDs, including STGD-like disease, RP, and CRD23. In our study, we observed PROM1 deletions in 3 RP families and one family with STGD-like phenotypes. In families FM157, FM297, and FM289, compound heterozygous and homozygous deletions in PROM1 were detected in probands, whereas their relatives with normal phenotype each carried a single heterozygous variant. Combining family history, AR mode of inheritance, clinical manifestation consistent with RP, as well as co-segregation analysis, the probands of the above three families were definitely diagnosed with RP. In FM112, the presence of a dominant heterozygous PROM1 deletion in three patients resulted in the onset of STGD-like disease. The fundus of the patients displayed atrophy of the retinal pigment epithelium (RPE) in the macular area of both eyes. Fundus photographs of the proband in FM112 showed a “beaten bronze” atrophic area of the macula and yellow pisciform flecks in the posterior pole of the retina. Meanwhile, FAF examination clearly showed the range of macular lesions (hypo-autofluorescence), with pisciform hyperfluorescent dots observed around the macula. SD-OCT revealed the loss of outer retinal structures in the macular area, accompanied by RPE atrophy and thinning (Supplementary Fig. 7). Interestingly, the same deletion (chr4:15992516–15997089) in the PROM1 gene was associated with two different clinical phenotypes (FM112, FM289, and FM297).
Moreover, due to overlapping phenotypes among various IRD conditions, accurate genetic diagnosis is crucial in refining clinical diagnoses for IRD patients9. For instance, patients in FM124 and FM130 were initially diagnosed with RP but were ultimately found to possess a hemizygous deletion in the CHM gene, the causative gene for choroideremia24. Advanced-stage choroideremia closely resembles end-stage RP, exhibiting similar chorioretinal atrophy and clinical symptoms, including night blindness and visual field constriction, with preserved central acuity. Genetic testing is imperative for a precise clinical diagnosis. Consequently, based on SV analysis of pathogenic genes, we conclusively diagnosed these two families with choroideremia.
In conclusion, our study highlights the potential of WGS to significantly enhance the diagnostic yield of IRDs and expand the mutational spectrum of known IRD-associated genes. The investigation of SVs and intronic variants holds substantial promise for the diagnosis and management of IRDs, facilitating personalized interventions for patients with these conditions.


留言 (0)