
記住我


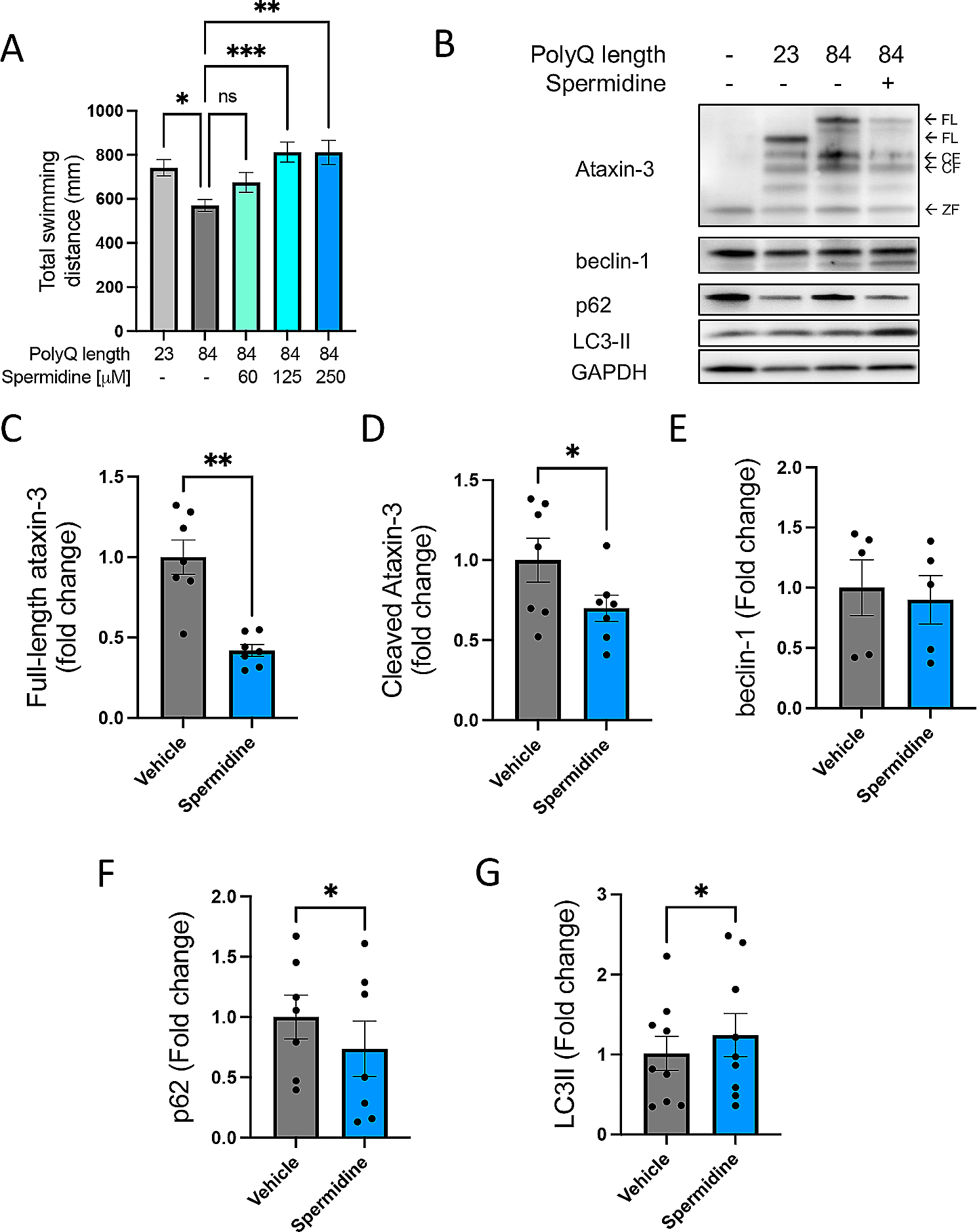


In this study, we conducted a comprehensive characterization of the mechanical profiles and mechanotransduction proteins within three distinct brain regions: the GW junction of the cortex, WM, and tissue extracted from the pons. The specific sampling locations within the brain are visually indicated in Fig. 1A.
To assess the mechanical behavior of the brain tissue, we examined stress-relaxation phenomena at a 7.5% strain. These data were effectively curve-fitted using a Zener model [2, 20], characterized by its spring term (\(\alpha\)), decay term (b), and equilibrium stress (\(_\)). This viscoelastic model is described by the equation, \(_=_*\left(a^ \right) + _\). Our analysis revealed significant variability in both the spring term and equilibrium stress within each brain region, as well as variations across different regions when individual donors were examined (Fig. 1B). Notably, significant differences in equilibrium stress were observed between the GW junction and the corona radiata, as well as between the GW junction and the pons in donor 3 (Fig. 1B). Furthermore, in donor 4, the equilibrium stress between the GW junction and the pons was found to be significantly higher (Fig. 1B).
When data from all donors were aggregated (data not shown), it became evident that the GW junction exhibited a larger spring term and equilibrium stress term compared to the pons, with statistical significance (p < 0.05). Utilizing a quasi-static compression model [20], we generated tissue compression data and subsequently calculated the elastic modulus. Comparative analysis of the elastic modulus at 2.5% strain under quasi-static compression revealed that GW junctions exhibited greater stiffness in comparison to regions such as the pons and WM (Fig. 1C).
Given the observed variability in mechanical properties across different brain regions, as depicted in Fig. 1B and C, our study aimed to identify the pivotal mechanosensors characterizing each region. Considering the clinical profiles of the donors, we conducted both pooled and individualized profiling of cellular mechanotransduction markers, including YAP, pYAP (phosphorylated YAP), β-catenin, and the mechanosensitive protein Piezo1. Notably, Piezo1 channels are transmembrane proteins highly expressed in neurons and glia [21, 22]. They play a crucial role in sensing changes in microenvironmental stiffness and transmitting these mechanical cues into intracellular signals [17, 23]. To investigate the distribution of Piezo1-positive cells across different brain regions, we conducted staining and quantification (Fig. 1D and E). Interestingly, while the other brain regions exhibited a relatively consistent range of 40–50% Piezo1-positive cells, the pons displayed a significantly higher proportion in three out of four donors (Figs. 1E and 2F). These consistent patterns of Piezo1 expression across brain regions were further confirmed using capillary electrophoresis and immunoblotting (Fig. 2A).
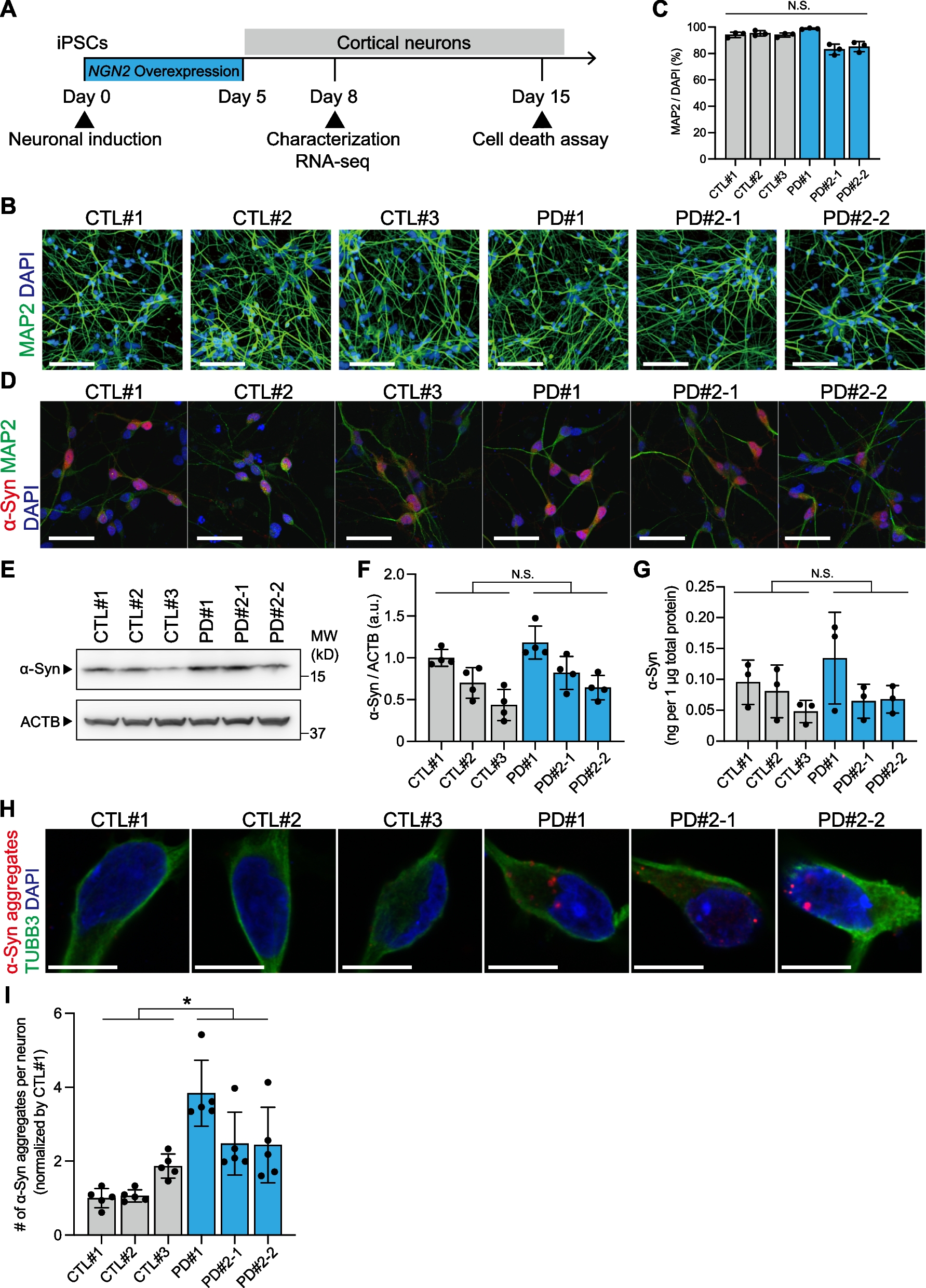
Fig. 2
Piezo1 expression profiling, cellular mechanotransduction, and biomechanical correlation analysis across brain regions of different donors. The relative abundance of (A) Piezo1, (B) YAP, (C) pYAP, and (D) β-catenin in the regions of WM (magenta), GW junction (purple), and pons (green) for individual donors (D1-4). (E, top left) Spearman correlation between Piezo1 relative fluorescence units (RFU) and stiffness; GW Junction (orange) ns, WM (blue) r = -0.5341, p < 0.05, pons (green) ns. (E, top right) Correlation between Piezo1 regional RFU and spring term (\(\alpha\)); GW junction r= 0.8791, and WM r = -0.5341, p < 0.05. (E, bottom left) Correlation between Piezo1 regional RFU and decay term (b); WM r = -0.6758, and pons r = 0.7692, p < 0.05. (E, bottom right) Correlation between Piezo1 regional RFU and equilibrium stress term (\(_)\); WM r = -0.8571, pons r = 0.7198, p < 0.05. (F) Comparison of protein expressed based on brain region for all donors, *p < 0.05
We explored the expression of Piezo1 downstream effectors, including YAP and its phosphorylated (inactive) isoform pYAP (Fig. 2B and C, and 2F). Notably, GW junction exhibited high YAP expression levels, with relatively high abundance in Donor 3. Moreover, the distribution of β-catenin expression showed unique patterns among regions across different donors and was notably increased in the GW junction of Donor 3 (Fig. 2D). Remarkably, our analysis revealed a significant correlation between Piezo1 expression and the stiffness of WM (Fig. 2E, top left). Additionally, viscoelastic parameters such as spring term displayed a positive correlation with Piezo1 expression levels in the GW junction while the correlation with WM showed a negative correlation (Fig. 2E, top right). The decay phenomenon (parameter ‘b’) and equilibrium stress (\(_)\) (Fig. 2E, bottom left and bottom right respectively) appeared to be significantly correlated with Piezo1 intensity in WM and the pons. Furthermore, donor-specific differences in YAP and pYAP expression have been observed. Thus, the biomechanical characteristics associated with Piezo1 vary depending on the specific region and individual variations among donors.
To gain a more comprehensive understanding of brain mechanotransduction and identify potential mechanosensors beyond Piezo1, we analyzed the expression profiles of Piezo2 and TMEM150C in three distinct brain regions: WM, GW junction, and the pons (Fig. 3). Similar to Piezo1, we investigated Piezo2 and TMEM150C from both expression level (relative fluorescence units, RFU) and the percentage of positive cell population.
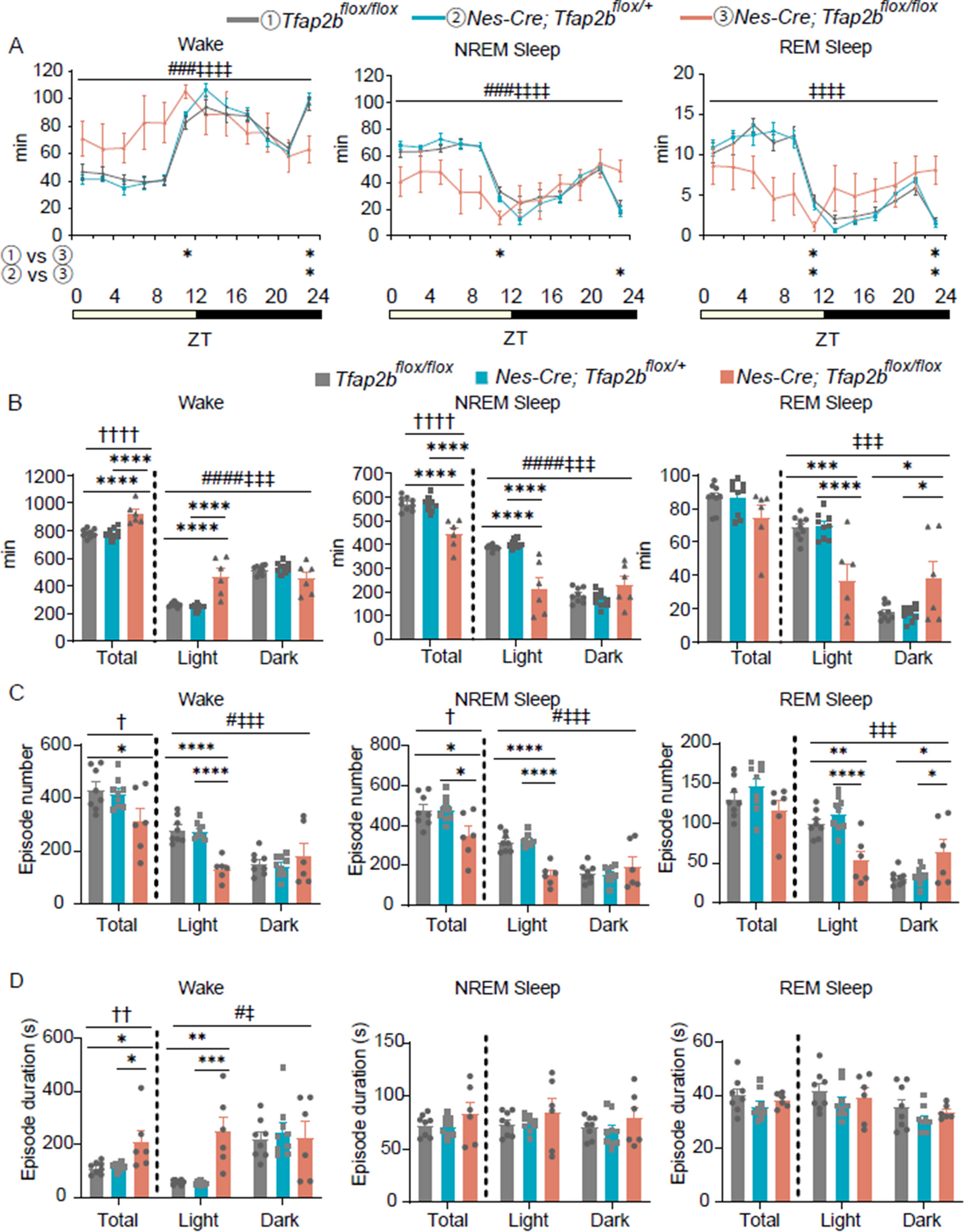
Fig. 3
The correlation of Piezo2/TMEM150C relative fluorescence units and percentage of positive cell population to brain mechanical properties. Percentage of Piezo2 positive cells (A) and TMEM150C positive cells (B) across different brain regions (*p < 0.05, **p < 0.01, ***p < 0.001). Spearman correlation between relative mechanosensor RFU and stiffness for Piezo2 (C) and TMEM150C (D) shows a significant correlation between Piezo 2 RFU and stiffness in WM only (r = 0.5516, p < 0.05). Spearman correlation between mechanosensor RFU and spring term for Piezo2 (E) and TMEM150C (F) with significant correlation in the Pons for Piezo2 RFU-spring term (r = -0.5602, p < 0.05) and TMEM150C RFU-spring term (r = -0.6429, p < 0.05). Spearman correlation between mechanosensor RFU and decay term for Piezo2 (G) & TMEM150C (H) showing no significant correlations across all brain regions. Spearman correlation between RFU and equilibrium stress for Piezo2 (I) and TMEM150C (J) where a significant correlation exists between TMEM150C RFU and equilibrium stress in white matter only (r = 0.5604, p < 0.05)
Our results revealed intriguing patterns of expression for both Piezo2 and TMEM150C across these brain regions. In Fig. 3A, we observed diverse expression patterns associated with Piezo2 expression in these regions. Notably, in Donor 1 (D1), there were no significant differences in Piezo2 expression levels between WM, GW junction, and pons regions. This suggests that Piezo2 may fulfill a similar mechanosensory role across these brain regions, indicating a shared mechanotransduction mechanism regulating brain activity. However, in Donor 2 (D2), Donor 3 (D3), and Donor 4 (D4), Piezo2 expression exhibited significant disparities. Specifically, in D2, WM displayed significantly higher Piezo2 expression compared to the pons, while in D3, Piezo2 exhibited higher expression in WM than the GW junction. In D4, the WM region demonstrated higher Piezo2 expression compared to both the GW junction and the pons (Fig. 3A).
The expression profile of TMEM150C, another candidate mechanosensor, presented intriguing findings in Fig. 3B. In D1 and D4, there were no significant differences in TMEM150C expression levels between WM, GW junction, and the pons. In contrast, D2 and D3 displayed a distinct elevation of TMEM150C expression in the WM compared to the GW junction or pons regions (Fig. 3B). This might suggest variations in TMEM150C across these regions that differ among individual donors.
To further explore the relationship between mechanical properties and Piezo2/TMEM150C (both expression level and percentage of positive cell population) across brain regions, we conducted correlation analyses similar to what was done for Piezo1. Notably, we observed significant positive correlations between stiffness and Piezo2 RFU in the WM (r = 0.5516, p < 0.05). Regarding the spring term (\(\alpha\)), we identified a significant correlation with Piezo2 RFU in the pons (r = -0.5604, p < 0.05) and TMEM150C RFU in the pons (r = -0.6429, p < 0.05). Finally, a correlation was observed between TMEM150C RFU and equilibrium stress in the WM only (r = 0.5604, p < 0.05) (Fig. 3C-J).
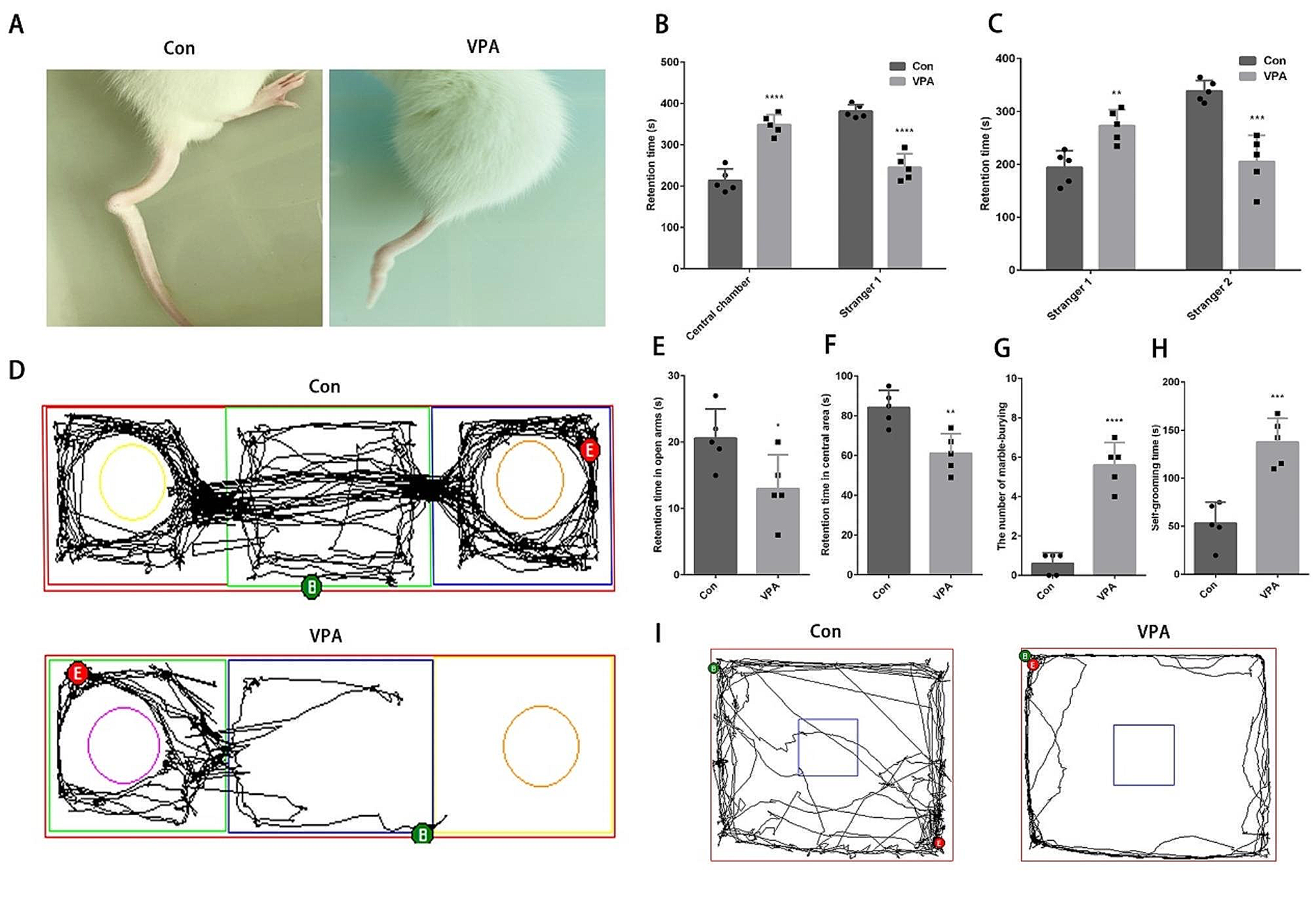
Hence, we identified Piezo1, Piezo2, and TMEM150C as mechanosensors in WM, GW junction and the pons through the correlation analyses linking region-specific expression (expression levels and the percentage of positive cell lines) with corresponding biomechanical characteristics as shown in Fig. 1E (Piezo1), Fig. 3A (Piezo2), and Fig. 3B (TMEM150C). We further sought to explore the three-dimensional localization of Piezo1, Piezo2, and TMEM150C in WM, GW junction, and the pons. An anatomical legend in Fig. 4E provides a visual reference for the location of each tissue region in a coronal view, illustrating the dimensional layering of the tissue sections, followed by a sagittal view illustrating the spatial arrangement of each tissue region. Our results demonstrated that the staining patterns of Piezo1, Piezo2, and TMEM150C remained consistent across different vertical planes within the tissue sections, as shown in Fig. 4A-D.
Fig. 4
Three-dimensional expression profile of Piezo1, Piezo2, TMEM150C across brain regions (WM, GW Junction, Pons) from four donors. (A-D) The top three rows represent the expression profile of Piezo1; the middle three rows represent the expression profile of Piezo2; the bottom three rows represent the expression profile of TMEM150C. (A-D) Columns represent the spatial position of each field of view showing the top, middle, and bottom sections of tissue in each brain region. Depth measurements for each field of view are labeled in the top right corner of each image (A-D). (A) expression analysis of and mechanosensor Piezo1, Piezo2, TMEM150C across brain regions (WM, GW Junction, Pons), (B) Donor 2 IF images for each brain region and mechanosensor, (C) Donor 3 IF images for each brain region and mechanosensor, (D) IF images for each brain region and mechanosensor Donor 4. (E) Anatomical legend for each brain region showing a coronal cross section (top) with an example of the spatial arrangement of each tissue layer for a particular brain region; the sagittal section (bottom) shows the spatial arrangement of each brain region. Scale bar = 50\(\mu\)m
留言 (0)