
記住我


To explore neural-related DNA hypermethylation in different cancers, we first performed differential DNA methylation analysis on the 144,004 probes that are located in the promoter region (29.7% of the total Infinium Methylation 450k Bead Array probes), corresponding to 21,089 Ensembl gene ids. Throughout this manuscript, we first explore results in colon adenocarcinoma (COAD), followed by a pan cancer analysis. We identified 11,659 hypermethylated promoter probes in COAD, resulting in 2624 genes with at least one hypermethylated probe in their promoter (logFC > 1, adjusted P < 0.05, and mean difference in B-value > 0.1). Additionally, all tested cancer types featured hypermethylation in about 1700 to 3400 genes, except for thyroid cancer that showed as little as 207 hypermethylated genes (Table 1). Figure 1 shows the top 10 enriched Gene Ontology (GO) terms and the top affected pathways in the 2624 hypermethylated genes observed in COAD compared to normal colon tissue. The lists of significantly differentially methylated probes in all the tested cancer types are provided in Additional file 1: Significant Limma output per cancer type.xlsx.
Table 1 Enrichment of neural genes in hyper- and hypomethylated genes for solid cancersFig. 1
Enrichment of neural terms in hypermethylated genes in cancer. Panel A shows an enrichment map plot of GO enriched terms for hypermethylated genes in COAD. Panel B shows a tree plot of the top Reactome pathways enriched for hypermethylated genes in COAD
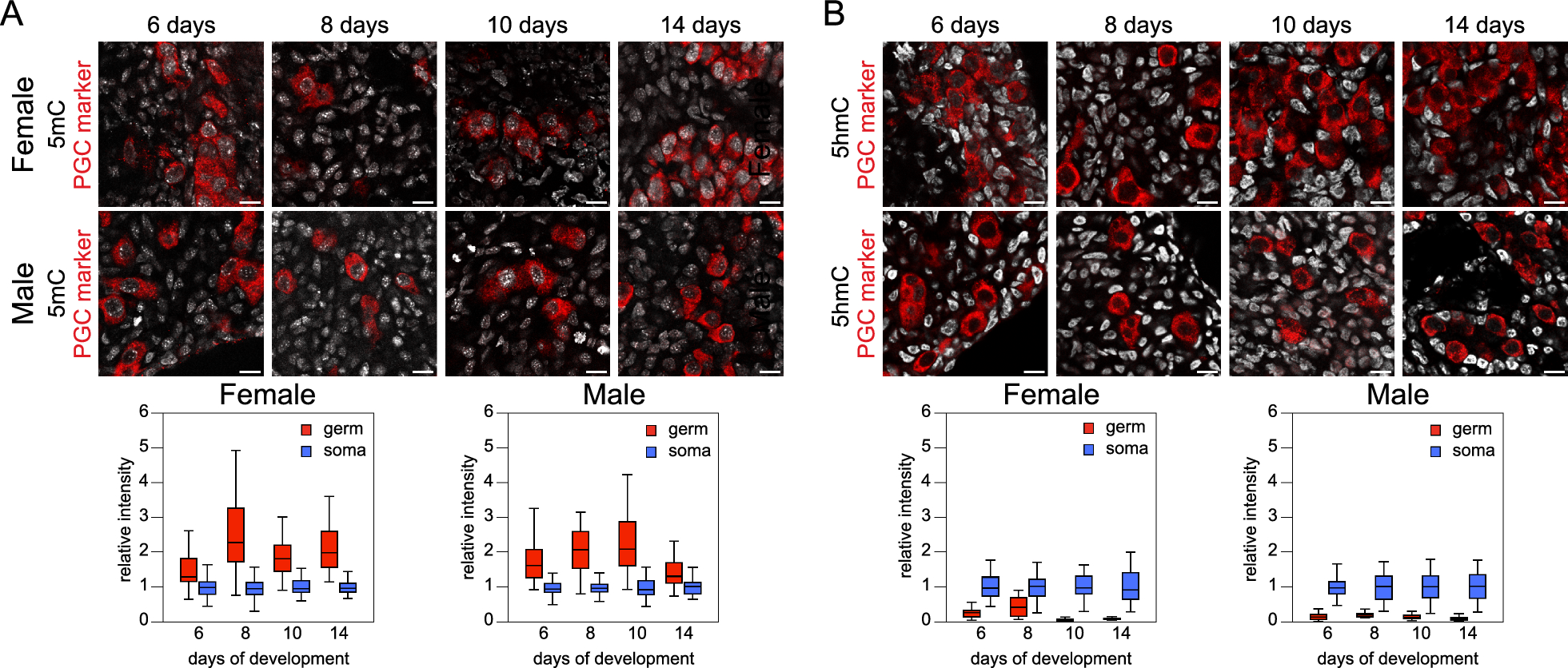
As described before for COAD [14], hypermethylated genes are enriched with neural-related genes (based on GO, see “Methods”): 38.5% of all promoter hypermethylated genes (n = 2624) were flagged as neural-related in COAD, whereas only 18.6% would be expected based on the percentage of neural-related genes on the Infinium array (further referred to as “Background”). Strikingly, this enrichment of neural-related genes amongst hypermethylated genes is consistently present in all cancer types (25.4–41.8%; all FDR-adjusted P values < 1.0E−4; Table 1 and Fig. 2), including thyroid cancer. Similar enrichment in all cancer types was found also when only considering DNA methylation in CpG islands into account (Additional file 2: Figures: S1 and S2, Table S1). As for hypomethylation, a similar enrichment of neural-related genes is only seen in cholangiocarcinoma and additionally, 7 out of 16 cancer types even show a derichment of neural-related genes (FDR-adjusted P values < 0.05; Table 1 and Fig. 2). We therefore concluded that the relationship with neural-related genes is particularly associated with hypermethylation.
Fig. 2
Percentage of neural-related hyper- and hypomethylated genes in different solid cancers. Percentage of neural-related hypermethylated (left) and hypomethylated (right) genes for all analyzed solid cancer types. A paired two-tailed t-test was used to compare percentages of neural-related hypomethylated and hypermethylated genes
Similarly, Reactome pathway enrichment analysis revealed that the pathway “Neuronal System” (r-has-112316) is enriched in all cancer types. Synaptic and ion channel-related pathways are also commonly enriched (15/16 cancer types). In addition, GO analysis resulted in several neural-related terms to be commonly enriched (15/16 cancer types): cellular component (CC) terms such as axonal components (GO:0043679, GO:0150034) and synaptic membrane components (e.g., GO:0099240, GO:0099572, GO:0097060); biological process (BP) terms such as dopaminergic neuronal differentiation (GO:0071542) and regulation of nervous system development (GO:0051960); and lastly the molecular function (MF) term: postsynaptic neurotransmitter receptor activity (GO: 0098960; Additional file 2: Table S2). Altogether, this analysis showed particular hypermethylation enrichment of neural-related and developmental genes during tumorigenesis. Especially, the neurotransmitters’ dopamine, norepinephrine and serotonin, as well as GABA and glutamate receptor genes were often affected by DNA hypermethylation. A list of all enriched GO terms per cancer type is provided in Additional file 1: GO outcome.rar.
The neural-related hypermethylation fingerprint is relevant for multiple hallmarks of cancer and is already present in premalignant tissueSubsequently, we evaluated to which extent neural-related hypermethylated genes were relevant throughout the hallmarks of cancer. Overlap with gene sets for known hallmarks of cancer [15] showed significant enrichment for neural-related genes amongst differentially hypermethylated genes (HMGs) compared to non-HMGs (FDR < 0.05) for 7 out of 10 studied hallmarks of cancer (Fig. 3, not significant for growth suppressor evasion, angiogenesis induction and tumor promoting inflammation) These results demonstrate that hypermethylation of neural-related genes is relevant for multiple aspects of carcinogenesis.
Fig. 3
Distribution of neural-related and non-neural-related genes in different hallmarks of cancer. Distribution of neural-related and non-neural-related genes for non-hypermethylated genes (left) and hypermethylated genes (right) for the different hallmarks of cancer
After establishment of the neural-related fingerprint in a variety of solid cancers, we evaluated whether this signature appears during early carcinogenesis by analyzing premalignant colon and bile duct samples and corresponding controls. 978 (35.8%) HMGs were found to be neural-related in premalignant colon adenoma samples, a significant enrichment compared to the background (Chi-squared test, P value < 0.0001). Also, for premalignant bile duct samples a similar enrichment was found, 1316 (32.2%) HMGs were neural-related genes (Chi-squared test, P-value < 0.0001). Despite enrichment of neural-related hypermethylated genes in premalignant tissue compared to normal tissue, there is still a significant increase in the enrichment when considering actual cancer tissue—35.8% vs 38.9% in colon cancer (P = 0.024) and 32.6% vs 37.0% cholangiocarcinoma (P = 0.0003). Of interest, neural genes being hypermethylated in premalignant tissue were typically also hypermethylated in solid cancer (data not shown). These results hint at a progressive mechanism, although a technical origin (e.g., different degrees of tumor purity) cannot be excluded. Therefore, these results indicate that neural-related hypermethylation is a part of the early aberrant DNA methylation events during carcinogenesis that may further progress during tumor development.
The neural-related fingerprint cannot be explained by tumor purity, bivalency, and higher DNA methylation degrees of neural genesTumor purity refers to the percentage of cancer cells in the tumor microenvironment (TME). As the TME consists of several types of cells, including neural cells, enrichment of neural-related genes amongst hypermethylated genes could represent a biological bias, i.e., Due to a different fraction of neural cells in controls and cases. Therefore, we investigated whether the number of hypermethylated genes in a tumor depends on tumor purity for neural-related and non-neural-related genes. For COAD, samples that showed a higher tumor purity displayed a higher fraction of hypermethylated genes (both for neural-related and non-neural-related genes). Therefore, the enrichment of neural-related genes amongst hypermethylated genes is more likely to occur in cancer cells rather than in non-malignant TME cells (Additional file 2: Figure S3), though only microdissection or single cell experiments can provide conclusive proof.
Furthermore, hypermethylated neural-related genes did not feature higher (even slightly lower) methylation degrees in tumor samples than non-neural ones, indicating that the enrichment is not caused by the fact that hypermethylation of neural-related genes is easier to detect (Additional file 2: Figure S4).
Additionally, bivalent genes are known to be more susceptible to hypermethylation in cancer. Therefore, we evaluated the association between the bivalency status in embryonic stem cells (ESCs) and the neural-related status of genes. In conformity with what is known from the literature, we found that hypermethylation of promoter regions occurs preferentially in bivalent genes during carcinogenesis. Indeed, we find bivalent genes to be enriched for hypermethylation in COAD as well as pan cancer (Additional file 2: Figure S5 and Tables S3 and 4). However, hypermethylation of bivalent genes during carcinogenesis was present at very similar (even slightly lower) proportions for neural bivalent genes when compared to non-neural bivalent genes.
Cancer cells appear to hijack a neural-related stemness stateNext, the Ingenuity Pathway Analysis (IPA) software was used to perform an upstream regulators analysis on hypermethylated neural-related genes (vs. non-hypermethylated neural-related genes) to identify possible regulators. Results showed that the master neural specification regulator RE1-Silencing Transcription factor (REST) was the top activated regulator of neural-related promoter hypermethylated genes. This revealed a possible role for REST as mediator of the neural-related hypermethylation fingerprint observed in cancer cells. Figure 4A, B shows mechanistic networks of the top upstream regulator REST revealing predicted pathways that might be involved in this signature such as the WNT pathway. In contrast to REST, differentiation and cell fate specification regulators PTF1A [16], TLX3 [17] and NEUROG2 [18] were marked as inhibited in the IPA analysis, which is consistent with the hypermethylation and subsequent downregulation of neural-related genes (Additional file 2: Table S5). Together with the notion that epigenetics plays a major role in cellular differentiation, the combined enrichment for factors that regulate neural lineage differentiation and specification in this list of upstream regulators hints towards the hypothesis that the neural-related hypermethylation signature helps granting cancer cells the required neural stemness to thrive.
Fig. 4
Ingenuity Pathway Analysis (IPA) output summary. A An overview network of the major biological themes of the hypermethylated neural-related genes in COAD, and B a mechanistic network for the top predicted upstream regulator (REST) and HMGs in COAD
To test our hypothesis, we identified a set of putative neural differentiation-associated genes relying on publicly available expression data, where inducible pluripotent stem cells (iPSCs) were differentiated through neural stem cells into neurons [19]. By performing differential expression analysis on the control samples in their neural stem cell and differentiated neuronal stage, we identified putative neural differentiation-associated genes (n = 1083) as those featuring significantly higher expression in differentiated neurons compared to neural progenitor cells. The list of neural differentiation-associated genes is provided in Additional file 1: Neural differentiation-associated genes.xlsx. We found that 29.0% of the differentiation-associated genes were hypermethylated at their promoter regions, vs. 18.6% of all the tested genes in colon cancer (OR = 3.13, P value < 0.0001). Moreover, when only considering neural differentiation-associated genes that are also annotated as neural-related by GO terminology, the share of hypermethylated genes increases to 44.1% (177 genes out of 401, OR = 5.88, P value < 0.0001).
Interestingly, we found that up to 47.8% of these hypermethylated neural and differentiation-related genes, show a significantly reduced expression in colon cancer, versus only 31.3% for hypermethylated neural-related genes that were not overexpressed in differentiated neurons (OR = 2.04, P value < 0.0001, Fig. 5).
Fig. 5
Overview of differential expressed HMG, neural genes, and neural differentiation-associated genes. Fractions of significantly downregulated (DR), upregulated (UR) or non-differentially expressed (nonDE) genes hypermethylated in colon cancer, compared to neural-related and neural differentiation-associated genes
In summary, these results suggest that hypermethylation of neural differentiation genes might be an active mechanism to silence the expression of neural differentiation genes in colon cancer, thus granting cancer cells neural stemness properties.
留言 (0)