
記住我








We have previously shown that FfH2A.Z is essential in F. fujikuroi spp. [29], but its global distribution and function remains enigmatic. To get more insight into the role of FfH2A.Z, we approached chromatin immunoprecipitation followed by sequencing (ChIP-seq) as well as MNase-assisted digestion and subsequent sequencing (MNase-seq) in our reference strain F. fujikuroi IMI58289 (FfWT).
For this, FfWT was grown in synthetic liquid ICI supplemented with 6 mM glutamine as the sole nitrogen source for 3 days. For both assays, mycelia were cross-linked before harvest with formaldehyde. Unfortunately, the first ChIP analyses using a commercially available antibody against H2A.Z proved promiscuous and therefore, we proceeded with N- and C-terminal tagging of the native FfH2A.Z gene with a 3x-hemagglutinin- (HA) tag. The FfH2A.Z ortholog (FFUJ_01849) was identified earlier [29] using QuartetS [37]. Multiple sequence alignment with ClustalOmega [38] with already characterized orthologs of H2A.Z revealed 97.9%, 94.4%, and 72.9% amino acid identity with F. graminearum (FGSG_01627), N. crassa (NCU05347) and S. cerevisiae (YOL012C) H2A.Z/Htz1, respectively. Pairwise sequence alignment of FfH2A.Z and FfH2A using EMBOSSNeedle showed an overall amino acid identity of 53.1% and 64.1% similarity. The typical histone fold domain found in the histones H2A, H2B, and H3 as well as the extended C-terminal domain are present in FfH2A.Z (Fig. S1). Interestingly, even though F. fujikuroi and N. crassa H2A.Z share a sequence homology of 94.4% as well as an identical domain organization, loss of NcH2A.Z is not lethal in this species [30, 32].
The N- and C-terminal HA-tagged versions of FfH2A.Z, i.e., HA::FfH2A.Z (Fig. S2A) and FfH2A.Z::HA (Fig. S3A), respectively, were transformed into the native FfH2A.Z locus using a nourseothricin (natR) resistance cassette. Three independent transformants each, i.e., HA::FfH2A.Z_T2, T3, and T4 as well as FfH2A.Z::HA_T10, T18, and T21, were gained (Figs. S2B and S3B). Wild type-like FfH2A.Z gene expression was confirmed via real-time quantitative PCR (RT-qPCR, Figs. S2C and S3C). To verify if tagging of the native FfH2A.Z gene influenced fungal development, we assessed radial hyphal growth, asexual reproduction, and SM biosynthesis (Fig. S4A-C). Additionally, for the FfH2A.Z::HA strains, a pathogenicity assay was performed (Fig. S4 D). Neither radial hyphal growth, SM production nor pathogenicity were influenced in the FfH2A.Z::HA strains, while slightly aberrant growth and SM biosynthesis were observed for the HA::FfH2A.Z mutants. Conidiation for both strains was slightly decreased compared to FfWT. Western blot analysis of the N- and C-terminally tagged strains using an anti-HA- and an anti-H2A.Z-specific antibody revealed the successful tagging of FfH2A.Z for HA::FfH2A.Z_T2 and T3 as well as for all three independent FfH2A.Z::HA mutants (Fig. S4E). Since normal fungal growth was observed for the FfH2A.Z::HA strains, we arbitrarily chose FfH2A.Z::HA T10 for further analyses. FfWT and the FfH2A.Z::HA transformant were grown and harvested as described before and incubated with an anti-HA-specific antibody. Incubation of wild-type mycelia with the HA-antibody served as a negative control to exclude the cross-reactivity of the antibody. For MNase-seq, crosslinked mycelia were treated with MNaseI and mono-nucleosome units were isolated and sent for sequencing. ChIP-seq analysis revealed overall 5,140 +/- 500 (2 biological replicates) FfH2A.Z-specific peaks (counts per million reads (CPM) cutoff > 15), which are evenly distributed over all twelve chromosomes in FfWT (Fig. 1). The cutoff (CPM > 15) was determined by the height of all genome-wide detectable FfH2A.Z peaks falling into a 95% quantile.
Fig. 1
Global distribution of FfH2A.Z in F. fujikuroi (FfWT). FfH2A.Z::HA (ChIP-seq) was mapped against the F. fujikuroi genome to visualize the overall FfH2A.Z occupancy. Chromosomes are shown in dark gray, while centromeres are depicted in light gray. Genome-wide distribution of FfH2A.Z present in the FfWT strain is depicted in turquoise and absent from the control (light gray). CPM; counts per million reads, bp; base pair
From these 3,400 +/-300 peaks (\(\sim\)60%) are localized within the regions 200 bp upstream to 600 bp downstream of ATG sites. With this, we defined the FfH2A.Z occupation by the maximum of normalized coverage in the regions 200 bp upstream and 600 bp downstream surrounding the ATG-sites of genes (5’ region) being greater than 15 CPM in both replicates (Fig. S5). This resulted in 2,374 promoter regions that are associated with FfH2A.Z incorporation. Next, we used all 2,371 genes harboring FfH2A.Z in their 5’ region to generate nucleosome enrichment maps (Fig. 2A and B) based on the detection of the + 1-nucleosome, i.e., the first nucleosome downstream the nucleosome-depleted region (NDR) for each gene [39]. From a total of 14,813 annotated genes, for 11,579, the + 1-nucleosome could be detected. This resulted in 1,636 genomic regions where the + 1-nucleosome and FfH2A.Z could be detected. In general, a low nucleosome abundance upstream of annotated genes indicates the transcriptional start site (TSS). This genomic area is also referred to as NDR, a genomic site where DNA is highly accessible for general transcription factors and other chromatin-associated regulators which in turn can recruit RNA polymerase II (RNA Pol II) and initiate transcription of the underlying DNA sequence [40, 41]. The NDR is usually flanked by the − 1- and + 1-nucleosome, defining an upstream and downstream barrier for the transcriptional machinery [42]. In general, for F. fujikuroi, nucleosome units are prevalent at the + 1- and to a slightly lesser extent at the + 2- and + 3-nucleosome, as well as at the − 1-nucleosome. This is followed by nucleosome depletion over the gene body (Fig. 2A). Aligning the data from the nucleosome map to the FfH2A.Z-specific peaks from the ChIP-seq analysis, clearly showed that FfH2A.Z is present at the + 1- and + 2-nucleosomes but lacking from the − 1-nucleosomes as well as from gene bodies (Fig. 2B). This pattern is seemingly conserved also in other organisms, i.e., in S. cerevisiae, D. melanogaster, and A. thaliana, where H2A.Z consistently occupies + 1-nucleosome at distinct genetic loci in the direction of transcription [43,44,45]. For the parasite Plasmodium falciparum, PfH2A.Z shows enrichment in euchromatic intergenic regions, i.e., in the TSS, and spans 6–8 nucleosome units [46, 47]. Next, examining FfH2A.Z incorporation in nucleosome units in more detail, it is evident that FfH2A.Z is almost exclusively incorporated at the + 1-nucleosome. In case where FfH2A.Z is also incorporated at the − 1-nucleosome, it seems as if these nucleosomes function at the same time as the + 1-nucleosomes for another gene, i.e., bidirectional promoter (Fig. 2C), which is also true for S. cerevisiae [45]. However, analyses in human cells showed that both nucleosomes flanking the TSS are indeed occupied by H2A.Z [48].
Fig. 2
FfH2A.Z occupancy and nucleosome localization in F. fujikuroi (FfWT). (A) Nucleosome map of all genes in FfWT (1,636 gene regions). Gene regions (1 kilobase pair (kbp) to 5 kbp flanking the + 1-nucleosomes) are sorted by increasing gene length from top to bottom, arrows on the top and on the bottom indicate the position of the + 1-nucleosome and the nucleosome depleted regions, respectively. (B) Genes are in the same order as in A, the heatmap shows FfH2A.Z occupancy concerning the + 1-nucleosome. CPM; counts per million reads not log2 scaled, NDR; nucleosome-depleted region, bp; base pair. (C) Density plot of FfH2A.Z occupancy over the − 1-, + 1-, + 1-nucleosomes from bidirectional promoters and all other nucleosomes. The threshold was determined based on the height of all genome-wide detectable peaks falling into a 95% quantile. CPM; counts per million reads
To conclude, the histone variant FfH2A.Z is predominantly incorporated non-randomly at the + 1-nucleosome on a genome-wide level in F. fujikuroi. Here, a larger fraction of FfH2A.Z is incorporated at the + 1-nucleosome of bidirectional promoters, which may serve simultaneously as a -1-nucleosome for the opposite gene.
FfH2A.Z is enriched at the 5’ region of actively transcribed genes in F. fujikuroiThe histone variant H2A.Z assumes pivotal roles in shaping the transcriptional landscape as already reported for several organisms [21, 26]. As we were curious to study the role of FfH2A.Z in transcriptional regulation also in F. fujikuroi in more detail, we first approached down-regulation of FfH2A.Z gene expression via the dosage-dependent tetracycline-responsive knock-down system TetOff (TetOff::FfH2A.Z) [49] in combination with an in situ constitutive overexpression of FfH2A.Z (OE::FfH2A.Z) in FfWT.
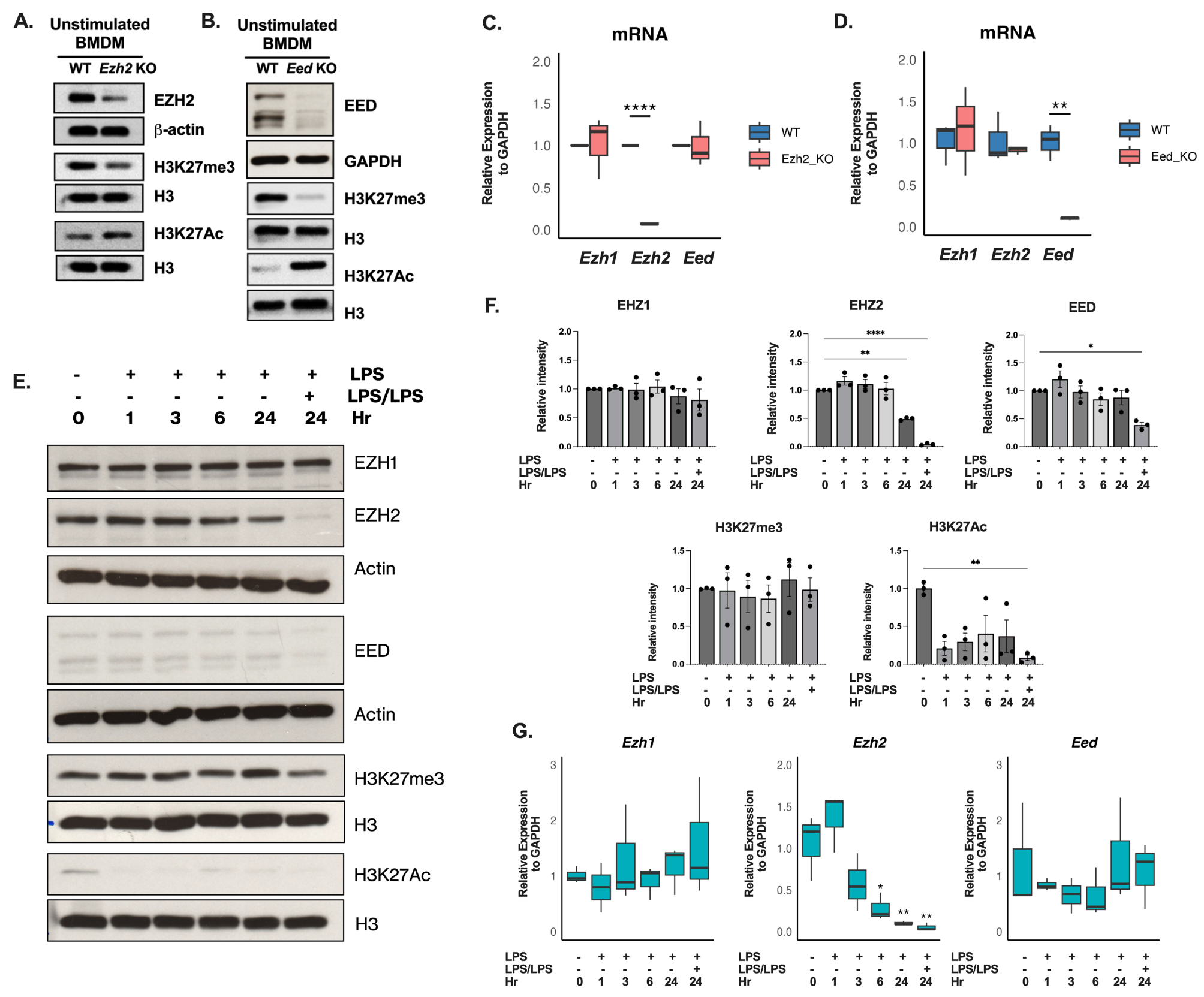
Strain generation of the knock-down and overexpression of FfH2A.Z was performed via homologous integration of the respective constructs into the native FfH2A.Z locus using a hygromycin resistance (hygR) cassette for the positive selection of transformants (Figs. S6A and S7A). Three independent FfH2A.Z knock-down transformants were gained, i.e., TetOff::FfH2A.Z_T1, T2, and T6 (Fig. S6B), while several transformants harboring the FfH2A.Z overexpression construct were obtained and arbitrarily three of them (OE::FfH2A.Z_T1, T3 and T4, Fig. S7B) were chosen for further analysis. Successful knock-down as well as overexpression of FfH2A.Z was verified via RT-qPCR and western blot analysis (Fig. 3A and B) using a commercially available anti-H2A.Z-specific antibody. To our surprise, FfH2A.Z expression in the knock-down strains under the addition of doxycycline hyclate (DOX) exceeded the expression of FfWT. However, western blot analysis clearly showed a nearly complete loss of FfH2A.Z on protein level under the same culture conditions. Later RNA-sequencing (RNA-seq) analysis of two independent knock-down mutants revealed wrong splicing patterns for FfH2A.Z transcripts, thus explaining increased transcription levels but lack of FfH2A.Z on protein level (Fig. S8). All transformants displayed a similar phenotype.
Fig. 3
Verification and characterization of OE::FfH2A.Z and TetOff::FfH2A.Z strains in Fusarium fujikuroi (FfWT). (A) Expressional analysis of the FfH2A.Z mutant strains OE::FfH2A.Z and TetOff::FfH2A.Z via RT-qPCR. For the FfH2A.Z depletion strain, FfH2A.Z expression was measured upon cultivation on CM supplemented with (50 µg/mL) and without (0 µg/mL) the inducing reagent doxycycline hyclate (DOX). The FfH2A.Z overexpression strain was cultivated on CM at 30 °C for 3 days in the dark. Gene expression of FfWT was arbitrarily set to 1. (B) Western blot analysis of FfWT, FfH2A.Z knock-down and overexpression mutant strains. Proteins were probed with an anti-H2A.Z-specific and an anti-H3 C-terminus-specific antibody. For quantification, a densitometric analysis was performed where FfWT was arbitrarily set to 1. (C) Assessment of radial hyphal growth of the inducible TetOff::FfH2A.Z strains and FfWT on CM using different concentrations of the inducing agent DOX. The medium was inoculated either with a mycelial plug (left panel) or 1,000 conidia each (right panel) and incubated for 4 days post inoculation at 30 °C in the dark. Experiments were performed in biological triplicates
Silencing via the TetOff system is dosage-dependent as shown earlier [49]. To determine the ideal DOX supplementation concentration, fungal growth on solid CM supplemented with different concentrations of DOX (0-100 µg/mL) was performed. Plates were point-inoculated either with freshly grown fungal mycelia or 10 µL of a 105 conidia/mL suspension of FfWT or TetOff::FfH2A.Z, respectively. Plates were incubated at 30 °C for 4 days. As depicted in Fig. 3C, reduced hyphal growth was observed for the knock-down strains already at 10 µg/mL DOX on plates inoculated either with mycelia or conidia. However, wild type-like “segmented” mycelia were observed next to crippled mycelia under the supplementation of nearly all tested DOX concentrations. The most stable growth of the mutant strains was observed under the addition of 100 µg/mL DOX but growth of FfWT was impeded drastically as well. To balance fungal growth and sufficient FfH2A.Z silencing, 50 µg/mL DOX was used for the induction of the TetOff::FfH2A.Z strains from now on.
Next, we performed RNA-seq with OE::FfH2A.Z, the TetOff::FfH2A.Z, and the respective wild-type strain using mycelia cultivated under the same conditions as used for ChIP- and MNase-Seq. Using a threshold of ≥ ± 1 for differential expression (log2) and a p-value < 0.01, 1,225 genes were upregulated and 1,455 genes were downregulated in the TetOff::FfH2A.Z mutant compared to FfWT, respectively (Table S1). Overall, 18% of the genome is deregulated, indicating neither an activating nor a repressing regulatory function of FfH2A.Z on a global scale. For the OE::FfH2A.Z strains, only 418 genes were up- and 180 downregulated, respectively (Table S1). Examining all genes that harbor FfH2A.Z in their 5’ region in more detail, we found that only 110 genes were up-, while 642 genes were downregulated by FfH2A.Z depletion. Thus, indicating that FfH2A.Z positively correlates with gene expression when incorporated near the TSS in F. fujikuroi (Fig. 4). These findings are similar as already shown for the ascomycete N. crassa [32], human CD4+ T cells [48, 50], and as indicated for Tetrahymena thermophila [51] where a positive correlation between H2A.Z incorporation and gene transcription was reported. This, however, stands in marked contrast to what has been described for other organisms such as S. cerevisiae [45], A. thaliana [36], D. melanogaster [52], and mammalian cells [53, 54], where a more dual role is ascribed to H2A.Z. Briefly, in S. cerevisiae ScH2A.Z marks the TSS of both silent and actively transcribed genes [45, 55] and the presence of ScH2A.Z at poorly transcribed genomic loci or even at heterochromatic regions [56, 57], where it is attributing to gene silencing, is evident. However, a recent study has shown that ScH2A.Z occupancy could not be unequivocally connected to either gene expression or gene silencing [45]. It appears that at this point, merely correlating H2A.Z occupancy to the transcriptional output is more complex than initially anticipated. However, it is noteworthy to mention that acetylation of H2A.Z (H2A.Zac) has been linked to positive gene transcription in several cell lines from chickens [58] to mammals [59, 60]. If this is also true for F. fujikuroi remains elusive at this point and is subject to further research. Hence, the regulatory role of H2A.Z in gene transcription remains somehow controversial depending on the organism.
Fig. 4
Global correlation between gene transcription and FfH2A.Z incorporation. The left-hand side (gray bars) shows barplot differential transcription of genes ordered by FfH2A.Z presence in the 5’ region and the level of differential transcription. The middle bar (red) indicates genes with no FfH2A.Z in their 5’ region, green indicates genes decorated with FfH2A.Z. The right-hand side shows a heatmap of transcription levels (RPKM; reads per kilobase pairs per million, normalized by log2) in the Fusarium fujikuroi wild type-strain (FfWT), the FfH2A.Z depletion strain (TetOff::FfH2A.Z), as well as the FfH2A.Z overexpression strain (OE::FfH2A.Z).
To study the function of genes affected by FfH2A.Z depletion in more detail, a Gene Ontology – Biological Processes (GO-BP)-based enrichment analysis of different subsets of genes was performed. Analysis of the genes occupied by FfH2A.Z revealed that 738 from 2,371 genes harboring FfH2A.Z are annotated by GO-BP and a total of 507 thereof are associated with: cellular macromolecule biosynthetic process (294), cellular macromolecule metabolic process (380), cellular nitrogen compound metabolic process (442), gene expression (361), nucleic acid metabolic process (362), regulation of biosynthetic process (214), regulation of nitrogen compound metabolic process (217) and regulation of transcription by RNA polymerase II (181) (Table S2). These results not only indicate that FfH2A.Z is crucial for general gene regulatory processes such as transcriptional control in F. fujikuroi, but also serve as a putative explanation for the observed secondary effects on the transcriptional levels in genes most likely not enriched in FfH2A.Z (Fig. 4). Next, to support this assumption, GO-BP annotation on both subsets of genes, i.e., positively or negatively affected by the FfH2A.Z depletion, was performed. Here, genes that are down-regulated in the TetOff::FfH2A.Z mutant strain indeed showed strong associations with amino acid activation (9), biosynthetic process (215), cellular biosynthetic process (199), cellular macromolecule biosynthetic process (147), cellular macromolecule metabolic process (178), cellular nitrogen compound biosynthetic process (161), gene expression (165), regulation of cellular biosynthetic process (97), regulation of metabolic process (103) and RNA metabolic process (140), which is in line with the results obtained in the GO-BP analysis performed on genes enriched in FfH2A.Z (Table S3). Hence, this further supports the assumption, that FfH2A.Z comprises vital roles in guarding the overall transcriptional balance in F. fujikuroi.
Lastly, when probing the gene set, which shows upregulation in the TetOff::FfH2A.Z mutant strain, GO-BP analysis revealed an enrichment in cellular response to DNA damage stimulus (17), chromosome organization (21), chromosome segregation (11), DNA metabolic process (23), establishment of chromosome localization (3), mitotic metaphase plate congression (3), positive regulation of chromosome segregation (3), response to mitotic spindle checkpoint signaling (2), tyrosine catabolic process indicating important. In general, H2A.Z is not only described to be involved in transcriptional control but also governs other fundamental roles, such as DNA repair and genome stability [61,62,63], which is also indicated by this analysis. Underrepresentation of FfH2A.Z leads to the upregulation of genes involved in chromosome stability and DNA repair. Interestingly, studies in S. cerevisiae revealed that loss of ScH2A.Z resulted in genetic instability caused by the accumulation of recombinogenic DNA damage through the non-coordinated action of the ATP-dependent remodeling complex SWR1 [64] and in consequence promoted genetic instability. It is tempting to hypothesize that genomic instability is the cause for the observed lethality upon loss of FfH2A.Z in F. fujikuroi [29]. However, this assumption needs further proof.
FfH2A.Z induces gene transcription regardless of the chromatin stateIn plants and higher eukaryotes, a correlation between histone marks, the histone variant H2A.Z, and gene transcription is evident [23, 33]. In detail, H2A.Z colocalization with histone marks at H3 and their impact on the transcriptional output are already well described for histone H3 lysine 4 trimethylation (H3K4me3) as well as histone H3 lysine 27 trimethylation (H3K27me3) in organisms such as the model plant A. thaliana [36], mESCs [34] or the ascomycete fungus N. crassa [32]. In general, H3K4me3 and H3K27me3 are of uttermost interest, since both marks are considered to act antagonistically regarding gene transcription. H3K4me3 mostly correlates with active gene transcription and is prominent in loosely packed chromatin (euchromatin), while H3K27me3 is deposited at transcriptionally inert regions with high nucleosome occupancy (facultative heterochromatin) [18, 20].
To test whether a similar co-localization pattern of H3K4me3 or H3K27me3 with the histone variant FfH2A.Z also exists in F. fujikuroi, FfWT mycelium was incubated with an anti-H3K4me3- and an anti-H3K27me3-specific antibody, respectively, followed by ChIP-seq. Obtained results show that overall 4,997 genes are enriched for H3K27me3 (max coverage upstream 100 bp downstream 1000 bp from ATG > 15 CPM), while 2,008 genes are decorated with H3K4me3 (max. coverage upstream 200 bp, downstream 600 bp from ATG > 20 CPM). In F. fujikuroi, H3K4me3, and H3K27me3 are largely exclusive (only 6 genes harbor both histone marks) on a genome-wide level (Fig. 5). Similar findings have been observed in Leptospheria maculans, Zymoseptoria tritici, Podospora anserina, and F. graminearum where both histone marks are either mutually or largely exclusive [65,66,67,68].
Fig. 5
Global distribution of H3K4me3, H3K27me3, and FfH2A.Z in F. fujikuroi (FfWT). All twelve FfWT chromosomes and their overall FfH2A.Z, H3K4me3, and H3K27me3 distribution. Chromosomes are shown in dark gray, while centromeres are depicted in light gray. FfH2A.Z occupancy is shown in turquoise, while decoration with H3K4me3 and H3K27me3 is shown in green and red, respectively. CPM; counts per million reads, bp; base pair
Comparison with the above described FfH2A.Z distribution data reveals that approximately 36% or 16% of all FfH2A.Z-containing nucleosomes are incorporated at genomic loci decorated with H3K27me3 or H3K4me3, respectively (Fig. 6A). If we now compare all genes decorated with either H3K27me3 (3688 genes) or H3K4me3 (1407 genes), which are not marked by FfH2A.Z (10,909 genes), we find 34% and 13% of facultative heterochromatin and trimethylated H3K4, respectively, in non-FfH2A.Z-marked genomic regions. These results already indicate that the presence of both, trimethylated H3K27 and H3K4 is largely independent of FfH2A.Z incorporation. Interestingly, for A. thaliana, O. sativa, P. falciparum, and mESCs a much higher co-occupancy rate between H3K4me3 and H2A.Z (\(\sim\)50 − 92%) was observed, compared to H3K27me3 (\(\sim\)20%) [34,35,36, 46, 47]. For the ascomycete N. crassa, the presence of NcH2A.Z is a prerequisite to establish H3K27me3 even if the histone variant and facultative heterochromatin are largely exclusive [32].
Fig. 6
Connection between histone mark distribution, FfH2A.Z, and their correlation with transcription in F. fujikuroi (FfWT). (A) Venn diagram of FfH2A.Z-marked genes versus H3K4me3- and H3K27me3-labeled genes, respectively. (B) Histogram of transcription levels in H3K4me3-/non-H3K4me3-enriched genes with and without the presence of the histone variant FfH2A.Z. (C) Histogram of transcriptional output in H3K27me3-/non-H3K27me3-enriched genes with and without FfH2A.Z. RPKM, reads per million per 1 kilobase (kb) normalized by log2
Correlating the transcriptional output to the H3K27me3- and H3K4me3-enriched regions shows, as expected, that H3K27me3 localizes to transcriptionally inert regions, while H3K4me3 is enriched in regions that are predominantly transcriptionally
留言 (0)