Animals
All experimental procedures were approved by the Institutional Animal Care and Use Committee at The University of North Carolina at Greensboro (protocol #19-006). Adult American bullfrogs (Lithobates catesbieanus) with no sex distinction weighing 100±15 g were acquired from Rana Ranch (Twin Falls, ID, USA) and housed in plastic tanks containing dechlorinated and aerated water in a 12/12 light/dark cycle. The animals were then randomly assigned to a control or an overwintering group, where hibernation conditions were simulated in the laboratory. Frogs in the control group were acclimated to the laboratory conditions for at least a week before the experiments. In this period, they were maintained at room temperature (23±2°C) with access to wet and dry areas and were fed with pelleted food provided by Rana Ranch once a week. Frogs in the overwintering group were placed in controlled temperature incubators (Thermo Fisher Scientific, Waltham, MA, USA), where the temperature was gradually reduced to 4°C over 10 days. Once at 4°C, screens were placed directly below the water level to impede access to the surface, and they were kept in this environment for 30 days before being used in experiments. Since the frogs do not eat in cold temperatures, overwintered animals were not fed. For the experiments, overwintered frogs were acclimated at room temperature for ~20 min before decapitation, and all the analyses proceeded at 22±2°C.
Dissection
The frogs were deeply anesthetized using 1 mL of isoflurane in a sealed container until loss of the toe-pinch reflex. They were then decapitated, and the head was immersed in the artificial cerebral spinal fluid (aCSF, composition in mM: 104 NaCl, 4 KCl, 1.4 MgCl2, 7.5 d-glucose, 1 NaH2PO4, 40 NaHCO3, 2.5 CaCl2, all purchased from Fischer Scientific, Waltham, MA, USA). This was bubbled with 1.5% CO2 and 98.5% O2 to oxygenate the brain while matching bullfrog arterial pH of ~7.85 at ~20°C [79, 80]. Decerebration was immediately performed, and the brainstem-spinal cord was carefully removed as described previously [37, 72, 81].
Tissue preparations
This study used several tissue preparations: intact, semi-intact, and brain slices. Below, we describe procedures for generating each type of preparation.
Motoneuron labeling
For all preparations involving motoneuron recordings, we labeled cell bodies for unambiguous identification of this cell type. Following dissection, the 4th branch of the vagus nerve complex was labeled bilaterally using a fluorescent dye (tetramethylrhodamine dextran 3000 MW; Invitrogen-Thermo Fisher, Waltham, MA, USA). This branch contains axons that mostly innervate the glottal dilator muscle, which is involved in lung ventilation in anuran amphibians [82, 83]. The nerve was pulled into a fire-polished pipette, and ~1μL of tetramethylrhodamine dextran 3000 MW was injected into the tip of the pipette in contact with the nerve for 1h on the first side and subsequently for 2h in opposite as previously described [27, 37, 72].
Semi-intact preparation
After labeling, the brainstem was horizontally sliced in a semi-intact preparation where only the vagal motoneurons are exposed, keeping the synaptic inputs of the native respiratory network. This preparation design and its output were previously described in detail [37]. Briefly, the brainstem was attached by agarose to an agar block keeping the most rostral side at a 45° angle, preventing it from being sliced. The vagal root was positioned over the angled edge (between 45 and 0°), maintaining the brainstem’s caudal part over the agar block’s horizontal side. This part of the brainstem was covered with agarose, and the block was glued to the vibratome plate (Technical Products International series 1000, St. Louis, MO, USA), where it was immediately immersed in oxygenated 4°C aCSF. In sequence, the dorsal part of the caudal brainstem was sliced at 200 μm until the bottom of the 4th ventricle was approached in the region between vagal and hypoglossal nerves. Then, 10- to 50-μm slices were cut to expose the region containing vagal motoneuron cell bodies, while the region containing neurons responsible for the lung rhythm generator was preserved [38, 84, 85]. The semi-intact preparation was pinned in a Sylgard-coated chamber for electrophysiology recordings (RC26G, Warner Instruments Holliston, MA, USA) and left to recover for 1 h before the recordings. In this period, the preparations were continuously superfused (~7mL/min) with aerated aCSF (98.5% O2 and1.5% CO2) using peristaltic pumps (Watson Marlow, Falmouth, CNL, UK) at room temperature (21°C±1).
Slices
In other experiments, the brainstem was sliced in cross sections to access labeled vagal motoneurons without rhythmic presynaptic input and neurons of the locus coeruleus (LC) [50, 86, 87]. For that, after dissection, the ventral side of the brainstem was glued to an agar block using super glue. The agar block was glued to the vibratome plate, and the vagal motoneurons were exposed by transversally slicing the vagal motor pool area at 300 μM (Burton and Santin, 2020; Santin et al., 2017; Zubov et al., 2021; Zubov et al., 2022). The midbrain was sliced at 400μM to access LC neurons [50, 87]. Additionally, the forebrain was dissected to access neurons of the pallium, the vertebrate precursor of the cortex [51, 88]. The pallium was dissected using the same method applied to access turtle cortical cells [89]. Briefly, after dissection, the forebrain was cut using an ophthalmic scissor dorsal to the lateral amygdala separating the pallium from the ventral regions. The pallium was then opened as a sheet, and dorsal pallium cells were used for the experiments [51].
ElectrophysiologyWhole brainstem—extracellular motor root and field potential recordings
The freshly dissected brainstem-spinal cord was pinned with the ventral side up in Sylgard (Dow Inc. Midland, MI, USA)-coated 6-mL Petri dishes where it was continuously superfused (~7mL/min) with aerated aCSF (98.5% O2 and 1.5% CO2) using peristaltic pumps (Watson Marlow, Falmouth, CNL, UK). All preparations were recorded at room temperature ~22°C. Borosilicate glass pipettes were pulled (Sutter Instruments, Novato, CA, USA) and manually adjusted in two sizes using sandpaper and fire polishing. A bigger size was used to ensure a tight seal around the vagal nerve root, and a smaller size was used to record from the lung generator cells. Rhythmic lung activity was recorded by an electrode placed in the area identified as the bullfrog’s lung rhythm generator [38, 39]. Extracellular signals from both regions were amplified (×1000) and filtered (low pass, 1000 Hz; high pass, 100 Hz) using an AM-Systems 1700 amplifier (Sequim, WA, USA). The signal was digitized using Powerlab 8/35 (ADInstruments, Dunedin, Otago, New Zealand), rectified, and integrated (100 ms τ) using the LabChart data acquisition system (ADInstruments, Dunedin, Otago, New Zealand).
Semi-intact preparation
The preparation was pinned in a Sylgard-coated recording chamber and transferred to the patch clamp set-up, where it was superfused with aCSF by gravity at a ~1–2-mL/min rate. The vagal nerve root was firstly identified at 4× magnification using a real-time imaging camera (Hamamatsu ORCA Flash 4.0LT sCMOS, Hamamatsu Photonics, Hamamatsu, SZ, Japan) and pulled into a fire-polished glass pipette suction electrode for extracellular recordings. After obtaining stable extracellular recordings to monitor network activity, labeled motoneurons were identified and used for patch-clamp experiments. The neurons were identified at 40× magnification and approached by glass pipettes (2–4 MΩ resistance) filled with a solution containing (in mM) 110-K-gluconate, 2 MgCl2, 10 HEPES, 1 Na2-ATP, 0.1 Na2-GTP, and 2.5 EGTA. The pipette was attached to a head stage (CV203BU) connected to an MP-285 micromanipulator and an MPC-200 controller (all Sutter Instruments, Novato, CA, USA). Positive pressure was applied to the tip of the pipette while approaching the cell and quickly removed, gentle negative pressure was used to form a >1GΩ, and the whole-cell access was obtained by breaking the seal with rapid negative pressure. After assuring stable access in the cell, voltage clamp was performed to verify the respiratory-related synaptic inputs (RRSIs) in neurons clamped at −66 mV. Membrane potential and respiratory-related firing behavior were measured in the current clamp. Firing ability was determined by a step protocol where the firing frequency-current (F-I) relationship was analyzed by injecting −150 to 1000 pA of current. All data were acquired in pClamp 11 software using Axopatch 200B amplifier and Axon Digidata 1550B digitizer (all from Molecular Devices, San Jose, CA, USA).
LC and pallium neurons
Slices containing LC cells and the pallium sheet were transferred to the recording chamber, stabilized using a nylon grid, and bathed with aCSF fed by gravity. After identifying and patching the cells as described above, membrane potential was measured in the current clamp, and the F-I relationship was determined in a step protocol. For LC neurons, the current injected for 0.5s ranged from −150 to 500 pA, and for pallium cells, −20 to 80 pA or until the cell achieves depolarization block.
Motoneurons in slices—evoked synaptic transmission
Slices containing labeled vagus motoneurons were transferred to a recording chamber and stabilized by a nylon grid. The motoneurons were identified (40×), and a bipolar tungsten stimulation electrode with tip separation of 250 μm (MicroProbes, Gaithersburg, MD, USA) was positioned ~120μm from the cell soma. The cell was then patched as described above and monitored in a voltage clamp, hold at −80mV. Nerve terminals in the slice were stimulated by 20-μs pulses every 2 s by an AM-Systems constant-voltage isolated stimulator (Sequim, WA, USA). The stimulus intensity was adjusted to produce a reliable synaptic response with the minimum voltage, which was monitored and adjusted if necessary for 2 min before starting experimental protocols to ensure the evoked current was stable.
Experimental proceduresWhole brainstem—extracellular motor root and field potential recordings
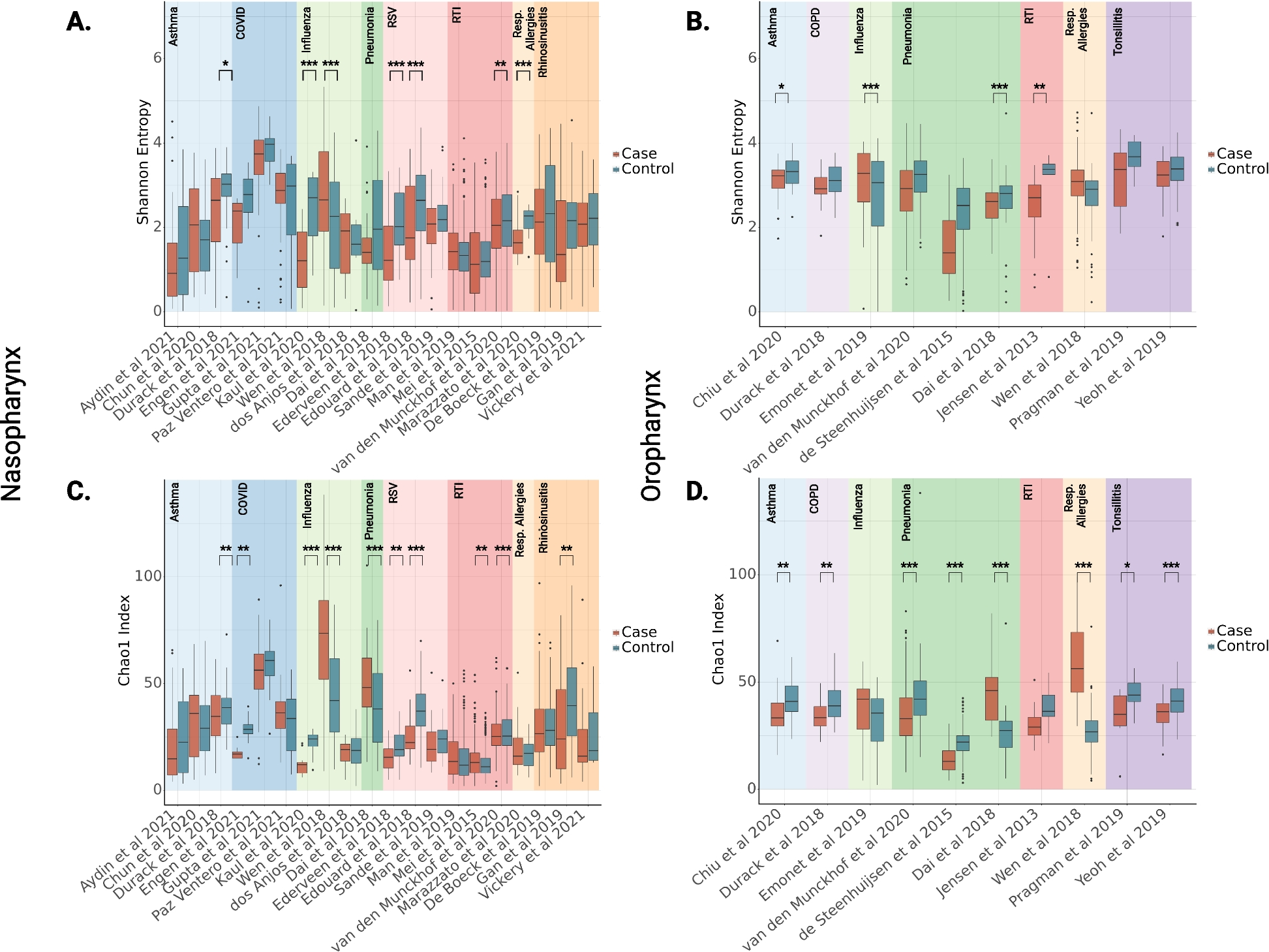
The output of whole brainstem preparation was monitored for 4h after decapitation to ensure a stable baseline for all recordings. Fictive breathing and activity of the lung generator were then recorded in baseline conditions for 10 min and throughout 4 h of hypoxia exposure, both for the controls and overwintered frogs.
Semi-intact preparation recordings
Motoneurons of controls and overwintered frogs had initial voltage and current measurements in baseline conditions (98.5% O2, 1.5% CO2) and, in sequence, were submitted to severe hypoxia, where aCSF was bubbled with 98.5% of N2 and 1.5% of CO2. Using this anoxic gas mixture, we previously observed ~2% of oxygen in the bath [27], which should result in tissue anoxia since the Po2 in the frog brainstem preparation has PO2 ~0 kPa near the brain surface and within the brain tissue at any depth when aCSF is gassed with a similar gas mixture [90]. While in hypoxia, voltage and current clamp protocols were alternated every 2 min. To investigate which cellular component was limiting network function, we compared the last measurements before network failure in control animals to 40 min of hypoxia in overwintered preparations in which all preparations still produced network activity. We used this approach because the respiratory network of control animals typically failed after a few minutes of hypoxia, and preparations of overwintered frogs keep working for several hours [27]. The time recorded in the motoneurons of overwintered preparations was limited by the inherent difficulty of maintaining long-term patch-clamp recordings in this preparation; 40 min represented the length of time that could be achieved in most neurons. The recording quality seemed to decrease over time only due to a technical limitation associated with maintaining quality patch seals and not as an effect of hypoxia since we could patch cells when the tissue was still hypoxic after the 40 min recording period (data not shown). Indeed, in an extreme example, we recorded one cell for the whole time until network failure after 2h of hypoxia exposure (Additional file 2: Fig. S1). Step currents were performed once in baseline conditions and once 10 min after the network failure in hypoxia for control animals. For overwintered animals, the second step proceeded after 40 min of hypoxia.
LC and pallium neurons
LC and pallium neurons had membrane potential verified, and step currents were injected in baseline conditions, which was followed by hypoxia exposure. Neurons were monitored in voltage and current clamp, and after 40 min of hypoxia, membrane potential and firing ability were analyzed again. Control and overwintered frogs were used in those experiments.
Motoneurons in slices—evoked synaptic transmission
To evaluate the hypoxia effect in the synaptic input per se, presynaptic terminals of vagal motoneurons isolated in slices were stimulated in baseline conditions. After assuring stable initial recordings in the voltage clamp, the slice was bathed with oxygenated aCSF containing 200nM TTX, 20 μM bicuculline, and 2 μM of strychnine to ensure that the evoked transmission had no influence from action potentials or inhibitory synaptic transmission. This solution was washed in for 2 min before the current was injected by the stimulator. After a stable current injection, the cell was recorded in voltage clamp while (a) perfused with aCSF+inhibitors, bubbled with 0% O2, and (b) monitored in an oxygenated solution for time control. Five stimulated postsynaptic currents (sEPSC) in baseline conditions were compared to the 5 last events in hypoxia or at 10 min of recording in time controls. Miniature excitatory postsynaptic current (mEPSC) was analyzed between stimuli at the same period sEPSC was analyzed.
Data analysis
Respiratory-related synaptic inputs, evoked synaptic current, and mEPSCs were recorded in voltage clamp, and their amplitude, width 50, peak area, and frequency were analyzed using the peak analysis function of LabChart (ADInstruments, Dunedin, Otago, New Zealand). The recordings were checked by eye to ensure the accuracy of the program in identifying the events. For mEPSC, a cutoff of 7pA was applied in amplitude. Input resistance was calculated using Ohm’s law (R=ΔV/ΔI) after recording the changes in voltage due to −100-pA current injection and was used to determine access quality. Membrane potential was determined in the current clamp in the period between respiratory bursts. The action potential threshold was considered at the last point before the action potential was triggered naturally by respiratory synaptic input. Firing frequency and the number of spikes triggered by RRSI included all spikes during a single event, which was analyzed for all events in a minute of current-clamp recording. Firing frequency in response to current was analyzed upon injection of 1000pA in vagal motoneurons (or until depolarization block), 200pA in LC neurons, and 60pA in pallium cells. Firing frequency in response to RRSI or current was calculated using the frequency function in the cyclic measurements function on LabChart (ADInstruments, Dunedin, Otago, New Zealand).
Statistical analysis
Data are raw values from individual experiments accompanied by mean. The hypoxic effect on RRSI’s and sEPSC’s peak area, width 50, and amplitude, as well as membrane potential, firing frequency, and mEPSC’s amplitude and frequency, were analyzed using paired t-test for both control and overwintered animals. Differences in activity time in the network/lung area and sEPSC were examined in a survival curve by the log-rank Mantel-Cox test. Statistical significance was accepted when p ≤ 0.05. Baseline values of control and overwintered frogs were compared using an unpaired t-test, and Welch’s correction was applied in case of different standard deviations. The number of cells and preparations (n) used for each experiment is indicated in the figures.








留言 (0)