1. IntroductionBecause of its being high in nutrients such as proteins, vitamins, trace elements, cellulose, and folic acid and having the characteristics of grain, fruit, and vegetable, sweet maize has received extensive attention in recent years [
1]. The energy provided by sweet maize is only 1/4 of that of kernel maize, and the sugar conversion rate is low; these make it widely loved by consumers. Due to insufficient accumulation of starch in the endosperm of sweet maize kernels, the seeds shrink after drying and lose water, becoming severely damaged. In addition, the high sugar content of the seeds and the large osmotic potential result in low seed vigor and emergence rates [
2].Due to the seed characteristics of sweet maize, seedlings of them are slender and weaker than those of common maize and are more suitable for shallow sowing. However, shallow-sowing cannot provide sufficient water to seeds, especially under drought and high-temperature conditions, a limitation which seriously affects sweet maize yield [
3]. A deeper soil environment has more water than a shallow sowing environment [
4] and can avoid chemical residues in shallow soil and damage caused to crops by wild animals [
5,
6], thereby stabilizing the primary form of the seed [
7,
8]. Further, due to the deeper distribution of crop roots, its drought resistance and lodging resistance can be enhanced [
9]. Deep-sowing inhibits the root length and root total surface area of maize at the three-leaf stage, but the root growth rate of maize after the five- to seven-leaf stage under deep-sowing condition is faster than that of the shallow-sowing condition [
10]. However, coupled with the germplasm characteristics of sweet maize itself, most of the existing sweet maize varieties have weak germination potential in the deep soil layer [
11]. Therefore, improving deep-sowing tolerance is the key to increasing sweet maize yield.Previous studies have shown that the mesocotyl is the key driver of seedling germination and establishment [
2,
6,
9], and maize varieties with longer mesocotyls have increased topsoil germination rates and higher seedling emergence [
12]. Furthermore, mesocotyl is the main area of maize to resist the invasion of soil-derived fungi and quickly respond to various abiotic stresses such as drought, low temperature, salt stress and waterlogging [
13,
14].There are many factors that affect the elongation of mesocotyl, among which plant hormones are vital internal regulators [
15]. Jasmonic acid (JA) may be involved in light-dependent regulation of mesocotyls [
16]. JA inhibits mesocotyl elongation, and mutants with impaired JA synthesis pathways have a longer mesocotyl phenotype [
17]. Ethylene inhibits gene expression in the JA biosynthesis pathway, reduces JA levels and promotes mesocotyl cell elongation [
18]. Brassinolide (BR) attenuates the inhibitory effect of light on mesocotyl by activating BZR1 to repress GATA2 transcription and reduce the accumulation of GATA2 [
19]. Indole acetic acid (IAA) mainly upregulates cell wall release activity, resulting in a continuous relaxation of the cell wall and mesocotyl elongation through cell growth [
20]. It has been reported that the IAA-binding protein ABP1 is actively involved in the elongation of the deep-seeding maize mesocotyl [
21].Research on the regulation of mesocotyl elongation by gibberellins (GAs) has been increasing in recent years. Exogenous GA3 application promotes the elongation of wheat internodes, rice mesocotyl and germinal sheath, and Arabidopsis hypocotyl and roots [
20]. GA regulates stalk elongation mainly by changing the orientation of cell microtubules, enhancing pectin methylation, and promoting cell elongation, which in turn promotes mesocotyl elongation [
22,
23]. In rice, exogenous GA3 can promote mesocotyl elongation, facilitate rapid seedling establishment and improve root early vigor, and GA combined with abscisic acid (ABA) can better promote mesocotyl elongation [
24,
25].GA20ox and GA3ox are the main sites for plant growth and development and environmental signals to regulate GA biosynthesis pathway, while GA2ox mainly regulates GA biological activity, inactivating active GA, thereby regulating the content of GAs in plants [
26,
27]. The ZmGA20ox1 gene overexpression line of maize showed increased biomass, plant height, stem length and longer leaf length under drought stress than WT [
28,
29]. The x rice GA20ox2 mutant sd1 showed significant decrease of plant height and GA content [
30]. Overexpression of AtGA20ox gene in kenaf significantly increased the content of active GAs, cellulose content and fiber length and quality [
31].Previous studies mainly focus on the molecular mechanism of rice mesocotyl development. Thus, the molecular mechanism of maize mesocotyl elongation needs further exploration. GA3 treatment of maize seeds is a simple and feasible method to improve deep-sowing tolerance during germination [
7]. In this study, to explore the role of GA in promoting mesocotyl elongation of deep-sowing sweet maize, we analyzed changes of phenotype and hormone content in sweet maize at different sowing depths. And we carried out the transcriptome data analysis and qRT-PCR data analysis. What’s more, ZmGA20ox1, ZmGA20ox4 and ZmGA20ox5 heterologous transformations to A. thaliana were also done. 2. Materials and Methods 2.1. Plant Materials and Culture Conditions
Seeds of sweet maize inbred line Ltx05 were used in this experiment and cultured in the greenhouse with a photoperiod of 16/8 h (day/night), a relative humidity of 65–70%, a light intensity of 600 μmol m−2 s−1 and a temperature of 28 ± 2/22 ± 2 °C (day/night).
2.2. Cultivation of Seedlings with Different Sowing Depths in Nutrient Soil
Seeds were cultured in nutrient soil at three sowing depths of 1, 5 and 10 cm with rectangular pots (51 × 37 × 14.5 cm, 30 seeds per pot) and watered every three days. The 15 days seedlings were used to determine the effect of different sowing depths on seedlings. Three replicates were performed for each group.
2.3. Treatment with Different Concentrations of GA3
The 1 cm and 10 cm sowing depths were chosen and separately treated with 0, 6, 10, 20 and 50 mg/L GA3 treatment solution for 7 days to determine the effect of GA3 on seedlings. Three replicates were performed for each group.
2.4. Treatment with GA3 and Gibberellin Inhibitor PP333
Seeds were surface-sterilized with 1% (v/v) sodium hypochlorite with constant shaking for 15 min and rinsed three times with distilled water before growth on sterilized filter paper in petri dishes (10 mm in diameter, 4 seeds per dish) for dark culture. The different groups were separately treated with 0, 20 and 50 mg/L GA3 and 200 mg/L PP333 (gibberellin inhibitor; polyconazole 15% wettable powder, WP), and the treatment solution was changed regularly and quantitatively every day for seven days. Ten replicates were performed for each group.
2.5. Measurement of Germination Rate, Mesocotyl Length and Root Surface Area
The germination rates of the different sowing depth groups were measured after 7 days. The mesocotyl and root systems of 15-day-old seedlings were rinsed with distilled water and dried with blotting paper. Mesocotyl length was measured with a ruler, and root morphology was imaged using a scanner (EPSON perfection V700 PHOTO, Dual Lens System, digital ICE technologies, Dhaka, Bangladesh).
2.6. Paraffin Sectioning MethodParaffin sectioning was performed according to our previous study [
32]. The mature zone of the mesocotyl parts was fixed with FAA fixative. Safranin and fast green were used to stain cut sections. After dehydration and mounting with neutral gum, the transect and slitting sections were observed using microscopic examination (Nikon Eclipse E100) and image acquisition analysis (Nikon DS-U3). 2.7. Determination of Endogenous Hormone ContentThe endogenous hormone contents of 1 and 10 cm sowing depth mesocotyls were determined by LC-MS (Liquid chromatography–mass spectrometry) with three replicates performed for each group [
33]. Quantitative samples were ground and added to an appropriate amount of internal standard (Olchemim/isoReag). Then, the powder was repeatedly extracted with acetonitrile solution in sequence. The supernatant was combined and fully reacted with TEA (triethylamine) and BPTAB (3-bromopropyl trimethylammonium bromide) and then blown dry with nitrogen. Finally, the samples were collected by filter membrane after re-solubilization with acetonitrile and later analyzed by LC-MS/MS. The MWDB (Metware Database) database was constructed based on the standards, and the data detected by mass spectrometry were analyzed qualitatively. 2.8. RNA Extraction and Transcriptomic Data Analyses
The total RNA from mesocotyl under 1 and 10 cm sowing depths at 15 days was extracted using TRIzol (Invitrogen, Waltham, MA, USA) and purified using the RNA easy Mini RNA kit (Qiagen, Hilden, Germany). High-throughput sequencing via an Illumina High-Seq2000 sequencing system was performed (Novogene Co., Ltd., Beijing, China), and genes with parameters of |log2(fold change)| > 1.5 and p < 0.05 were recognized as DEGs. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis were also performed.
2.9. qRT-PCR Analysis of DEGs in MesocotylsqRT-PCR was carried out in a Light Cycler 480 (Roche) using SYBR Premix Ex TaqTM (TaKaRa, Tokyo, Japan) according to the instructions. Mesocotyls at 7 and 15 days were used to measure the expression levels of candidate genes. The calculation of the relative expression value was performed according to the 2−ΔΔCt method [
34]. The qRT-PCR verification primers are shown in
Table S1. 2.10. Plasmid Construction and Heterologous Transformation of ARABIDOPSIS Col-0According to MaizeGDB (
https://www.maizegdb.org/, accessed on 2 December 2022), the total length of coding sequence of ZmGA20ox1, ZmGA20ox4 and ZmGA20ox5 of inbred line Ltx05 was cloned and verified by sequencing (
Supplementary Figure S1). The coding sequence of ZmGA20ox1, ZmGA20ox4 and ZmGA20ox5 without stop codon were inserted into pCAMBIA 3301 digested with Nco I. The constructs were introduced into the Agrobacterium tumefaciens strain GV3101 and then transformed into A. thaliana Col-0 by the Agrobacterium-mediated floral dip method [
35]. After 0.1% herbicide (v/v) screening for three consecutive generations, homozygous Col 35S:: ZmGA20ox1, 35S:: ZmGA20ox4 and 35S:: ZmGA20ox5 were harvested and identified by qRT-PCR. Line OE1-3, OE4-11 and OE5-5 with lowest expression level were selected as the control (relative level is one) to calculate the relative expression level of different transgenic strains. 2.11. The Hypocotyl Observation and Determination in Transgenic Lines
Seeds of three OE-ZmGA20ox1 (OE1-3, 1-10 and 1-9), OE-ZmGA20ox4 (OE4-11, 4-9 and 4-8) and OE-ZmGA20ox5 (OE5-5, 5-12 and 5-10) lines with low, medium, and high levels of ZmGA20ox1, ZmGA20ox4 and ZmGA20ox5 expression were placed on ½ MS medium (0.9 % agar) in square sterile culture boxes. Ten seeds per line were sown and ten biological replicates were performed. Homozygous seedlings cultivated for seven days were observed by dissecting microscope (Nikon, Tokyo, Japan) and hypocotyl length was measured.
2.12. Data Analysis
Experimental data were processed and analyzed with Duncan’s multiple range test or Student’s t test. All statistical analyses were carried out with SPSS v16.0.
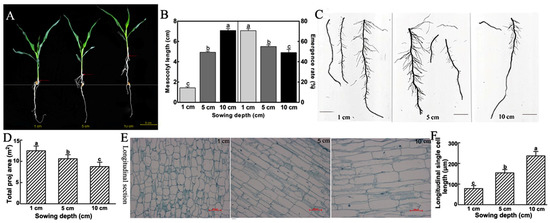
4. DiscussionAs a popular maize species, more and more studies have been carried out on the planting and growth of sweet maize. Studies showed that seedling emergence is directly related to mesocotyl elongation [
8,
13]. Our study showed that with the increase of sowing depth, the emergence rate of sweet maize inbred line Ltx05 decreased and the mesocotyls elongated, which indicates that the seedlings sown deeply completed the germination by promoting mesocotyl elongation. The results of the mesocotyl paraffin section showed that in the 1 and 10 cm sowing depth environment, the sweet maize germination was mainly completed by the longitudinal elongation of the mesocotyl cells. The root system data show that the shallow sowing seedling may ensure the supply of water and nutrients by increasing the number of root hairs and expanding the contact area with the soil (
Figure 1).Phytohormones play an important role in mesocotyl elongation and growth of sweet maize [
38]. Our determination of endogenous hormone content showed that deep-sowing can promote the synthesis of various plant hormones in the mesocotyl, especially IAA, GAs, ACC and tZ, so that the elongation of mesocotyl cells can be promoted. JA, instead, inhibited mesocotyl growth (
Figure 2). These are similar to the results reported in maize and rice [
16,
36]. Studies showed that IAA treatment promoted cell elongation of mesocotyls under deep-sowing environments [
20]. In rice seedlings, ethylene inhibits JA biosynthesis by decreasing the expression of gaoyao1 gene and other genes to reduce JA levels and enhances mesocotyl elongation [
18]. GA3 only or ethephon and GA3 treatment significantly promote the mesocotyl elongation of rice under 5 cm flooding depth stress [
39]. Results in this study and these previous studies indicated that ethylene in sweet maize Ltx05 played a positive role in mesocotyl elongation under deep-sowing stress. The GA53 content was only detected in the mesocotyl at a sowing depth of 10 cm, which may be due to the soil environment influence and pressure during deep-sowing on the regulation of GA53-related signaling pathways.Deep-sowing could reduce the emergence rate, but moderate GA3 treatment could alleviate the damage caused by this stress. GA3 had a better germination effect on seeds with low vigor. 20 mg/L GA3 was the best concentration to promote deep-sowing tolerance of sweet maize inbred line Ltx05 especially reflected in the improvement of germination rate (
Figure 3). The inhibitor PP333 could significantly inhibit the promoting effect of GA3 on seedlings and mesocotyl elongation (
Figure 4). The optimum concentration of GA3 for different maize varieties is also different [
7,
11].Major life activities depend on differential gene expression [
40]. Transcriptomic analysis revealed that sowing depth affected gene expression differences in mesocotyl. GO and KEGG analysis showed that deep-sowing stress affected the microtubule architecture, which ultimately affected the elongation of the mesocotyl and influenced the expression of oxidoreductase activity relative genes. Microtubules have been discovered to play an important role in mesocotyl elongation in rice [
41]. Cortical microtubules destabilization induces mesocotyl cell elongation, and this process is related to endogenous GA content [
42]. The hydrogen peroxide metabolic process also has been affected by sowing depth. A previous study showed that hydrogen peroxide pretreatment to seeds can improve deep-sowing stress tolerance of wheat [
43]. KEGG enrichment showed that a large number of DEGs (especially transcription factors) were enriched in plant hormone signal transduction, and these were accordance with the difference of hormone content index measured in this experiment. In rice, the expression level of OsTCP5 regulated by cytokinin and strigolactones (SL) is negatively correlated with the length of mesocotyl [
44]. The expression of ZmMYB59 decreases the germination and mesocotyl length of Nicotiana tabacum and rice [
45]. The difference in mesocotyl phenotype and the change in hormone content at different sowing depths must be the result of the synergistic effect of these hormones on mesocotyl growth, and the specific regulatory pathways need to be further studied [
13].Physiological mechanism and genes involved in mesocotyl elongation have been extensively studied [
46,
47,
48,
49]. However, there are relatively few studies on the molecular mechanism of GA in the process of mesocotyl elongation under deep-sowing stress [
34]. GA20ox played an important role in many life processes of the plant. The GA20ox1 gene is highly expressed in wheat stems [
50]. When GA20ox is overexpressed in rice, the plant exhibits internode elongation [
51]. The GA20ox1 gene is highly expressed in sorghum at the jointing stage [
52]. Arabidopsis transgenic lines overexpressing CsGA20ox1 evidently increase germination rate, rosettes area, leaf area and petiole length, and rosettes area and leaf area [
47]. In our qRT-PCR results, the expression levels of the GA20ox1, GA20ox4 and GA20ox5 genes in the mesocotyl of 7 days seedlings at the 10 cm sowing depth were higher than those in the 1 cm control and lower than those in the 1 cm control of 7 days seedlings. The GA20ox1, GA20ox4 and GA20ox5 heterologous transformation of arabidopsis significantly increased the hypocotyl length and germination rate. These results suggested that GA20ox1, GA20ox4 and GA20ox5 may promote the growth of mesocotyl in the early stage.






留言 (0)