Adipose tissue is a highly dynamic organ distributed throughout the body and can be classified according to the cellular composition of different sites and its unique anatomical localization [
36]. Different adipose tissues of the human body can be defined according to their location, size, cellular composition, and function and are mainly divided into white, brown, and beige adipose tissue [
37]. White adipose tissue (WAT) constitutes the largest proportion of adipose tissue in the body, seen in major organs and perivascular, abdominal, and subcutaneous sites. White adipose tissue, once thought to be morphologically and functionally unspecific, is now considered dynamic, plastic, and heterogeneous and is involved in a wide range of biological processes, including energy balance, glucose and lipid utilization, blood pressure control, and host defense [
38,
39,
40]. WAT stores excess energy in the form of triglycerides, and the increased accumulation of WAT, particularly in visceral depots, is a key determinant of cardiometabolic disorders, hypertension, and the relative risk of CVD12–17 [
41]. Beige and brown adipose tissue account for less than 5% of adult adipose tissue, but they play a key role in lipid metabolism, glucose metabolism, and body temperature maintenance [
42]. Brown adipose tissue is mainly stored in the interscapular region of infants and adults. Beige adipose tissue is a kind of white adipose tissue with scattered brown adipocytes, which has become of concern in recent years [
43,
44,
45]. The massive expansion and phenotypic remodeling of adipose tissue during obesity have differential effects on specific adipose tissue stocks, and the relationship between the type of adipose tissue and its effect on cardiovascular function becomes particularly evident in the context of the heterogeneous phenotype of pericardiovascular adipose tissue. The increase in white adipose tissue greatly contributes to vascular dysfunction and cardiovascular disease [
46,
47,
48,
49]. Thermogenic brown and beige adipose tissue are associated with more positive cardiovascular effects than white adipose tissue.Adipose tissue (AT) is a highly heterogeneous endocrine organ composed of mature adipocytes, preadipocytes, fibroblasts, endothelial cells, and a series of inflammatory leukocytes. The heterogeneity among adipose tissues from different anatomical sites results from their inherent differences in cellular and physiological properties, which are caused by developmental origin, adipogenic and proliferative capacity, glucose and lipid metabolism, insulin sensitivity, hormonal control, thermogenesis, and angiogenesis [
50,
51,
52]. Other factors that influence adipose tissue heterogeneity are genetic predisposition, sex, environment, and age. In the case of hyperplastic expansion of adipose tissue, these specific differences in adipose tissue translate into specific patterns of fat distribution and fat phenotypes, which are closely related to the risk of multiple metabolic diseases. The main functional cell type of AT is adipocytes. There are also obvious differences in the adipocyte morphology and function of different adipose tissue phenotypes. The main function of WAT is to promote a dynamic energy balance through the storage and release of fat (e.g., triglycerides) in response to nutrient intake and metabolic needs. Thus, white adipocytes contain a single large lipid droplet (monocular) and only a small amount of mitochondria [
53]. At normal temperatures, beige adipocytes have more white phenotypes, with large lipid droplets and a lower expression of thermogenic genes, but the cold response, β-adrenergic stimulation, or motor activation leads to a robust upregulation of the thermogenic program, a process often referred to as “browning” [
54]. Interestingly, since the epigenomic plasticity of beige adipocytes is temperature-dependent, there is also the ability to “whiten” in warm environments [
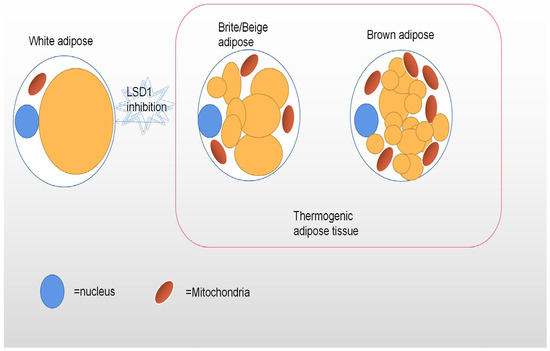
55]. Unlike white adipocytes, heat-producing adipocytes (brown and beige adipocytes), especially brown adipocytes, have multicompartment lipid droplets and contain a large number of dense mitochondrial inner membrane uncoupled protein-1 (UCP1) mitochondria (
Figure 1), which can oxidize fatty acid at a very fast rate [
56]. Their highly active cellular metabolism is the result of unique protein mechanisms that allow the thermogenesis of adipocytes to participate in the futile recycling of normally stored metabolites. White and thermogenous brown/beige adipocytes come from different precursors, but development of both brown and beige adipocytes depends on the transcriptional coregulatory protein PR domain 16 (PRDM16) [
57]. The thermogenic oxidation machinery of brown and beige adipocytes is regulated by transcription factors and cofactors, such as PR domain 16 (PRDM16) and the peroxisome proliferator-activating receptor γ coactivator-1 α (PGC-1 α), whose transcription factors bound to DNA cooperate to induce mitochondrial biogenesis and fatty acid oxidation of requiring heat production [
58,
59].






留言 (0)