1. IntroductionSnakebite envenoming is a life-threatening, morbidity-causing, World Health Organization (WHO)-listed neglected tropical disease [
1]. The populations at greatest risk of snakebite occupy the tropical and sub-tropical regions of the world, and the burden of disease is greatest in rural areas of South and Southeast Asia, sub-Saharan Africa, and Latin America [
2]. Collectively, it is estimated that perhaps as many as 1.8 million people are envenomed annually, of whom 90,000–138,000 die, and many more suffer life-long morbidity as the result of associated physical and mental trauma [
1,
2]. In Latin America, it is estimated that between 80,000 and 129,000 individuals are envenomed annually, though the true incidence may be greater due to extensive global underreporting of snakebite [
3].Pit vipers of the genus Bothrops (Viperidae: Crotalinae), often referred to as lanceheads or lancehead vipers, are of greatest medical importance in Central and South America, as they are responsible for causing the majority of cases of severe envenoming [
1,
3,
4]. Snakes of this genus inhabit a large geographical distribution, ranging from southern parts of Mexico in the north, to Argentina in the south, and also include Caribbean island populations, such as Bothrops lanceolatus on Martinique and B. caribbaeus on St. Lucia. The pathophysiology of envenomings by Bothrops spp. can be variable, but victims typically present with haemorrhage, hypotension, impaired blood coagulation, inflammation, acute renal damage, and/or local tissue damage, the latter of which can result in irreversible sequelae requiring surgical interventions such as the removal of necrotic flesh or amputation of extremities or digits [
3,
5,
6].Venom proteomic [
7,
8] and venom gland transcriptomic [
9,
10] studies have provided considerable insights into the venom composition, and thus the aetiological toxins responsible for causing envenoming pathology, of a large number of Bothrops species over many years. In line with many viperid species, toxins of the snake venom metalloproteinase (SVMP), snake venom serine proteinase (SVSP) and phospholipases A2 (PLA2) gene families are typically the most abundant components found in Bothrops venom [
11]. Although considerable inter-specific variation in venom composition exists within the genus, the SVMPs are typically the mostly abundant of these [
11] and this toxin family can represent as much as 75% of the total proteins present in venom, as observed in B. lanceolatus [
12]. SVMPs are enzymes with multiple functions that participate in both local and systemic effects of snakebite envenoming, mostly notably via the cleavage of basement membrane proteins of blood vessels resulting in haemorrhage, or via the activation or degradation of blood clotting factors resulting in coagulopathy, though these toxins can also interact with platelets and inflammatory mediators [
13]. The SVSP toxin family also contribute to venom induced coagulopathy, with many representatives classified as thrombin-like enzymes due to their fibrinogenolytic mode of action [
14], while PLA2s are more typically implicated in contributing to the severity of local envenoming effects, including myotoxicity, though they can exert anticoagulant activities via inhibition of platelet aggregation [
15,
16]. Abundances of these toxin types also vary across the genus, with PLA2s reaching a maximum abundance of 45% in B. asper, while SVSPs peak at 29% in southern populations of B. jararaca [
11,
17,
18]. Such inter-specific toxin family variations also extend to other minor venom components, including C-type lectins and disintegrins, while intra-specific sex- and population-based, along with ontogenetic, venom variation has also previously been described from this genus of snakes [
18,
19,
20,
21].The only specific treatments for snakebite envenoming are antivenoms, which consist of polyclonal antibodies sourced from venom hyper-immunised animal plasma/sera [
1], and their importance is highlighted by inclusion on the list of essential medicines published by the WHO (
https://www.who.int/publications/i/item/WHO-MHP-HPS-EML-2021.02 accessed on 13 September 2022). There are two main types of antivenom, namely monospecific (or monovalent) antivenom, which is generated against the venom of a single snake species, and polyspecific (or polyvalent) antivenom, which is made using the venom of multiple snake species as immunogens. There are advantages and disadvantages to both approaches: in general, for many parts of the world polyspecific antivenom is more advantageous because of the presence of multiple biting species and a lack of effective diagnostic tools; thus a single therapy that can be used for a particular region is desirable. However, due to only a proportion of the antibodies present in polyspecific antivenom being specific to the snake species responsible for a bite, typically higher therapeutic doses have to be administered to effect cure [
22], resulting in cost increases and a theoretical increased risk of adverse reactions. Moreover, despite the production of polyvalent antivenoms, venom variation renders many of these products with limited efficacy against different snake species not included in the immunising mixture, or even against different populations of the species whose venoms were used to generate the antivenom [
23,
24,
25,
26]. Sadly, this means that many people who suffer from snakebite envenoming do not have access to effective, specific therapeutics. The unpredictable nature of snake venom variation means that there is a strong rationale for testing the capability of existing antivenoms to inhibit the venom activities of snake species related to those used as immunogens. Evidence of paraspecific efficacy might increase the geographical utility of an antivenom, which could benefit patients by increasing access to effective treatment, while manufacturers may benefit by accessing new markets, so long as standards of safety and efficacy are upheld [
27].In this study, we explored the potential utility of three distinct antivenoms generated (at least partially) against medically important Bothrops spp. found in Central and South America at binding with and inhibiting the in vitro functional activities of seven distinct Bothrops venoms spanning a broad geographical distribution ranging from Costa Rica to Brazil (
Table 1). The antivenoms included the polyvalent anti-Bothrops Brazilian product Soro antibotrópico-crotálico, the trivalent anti-Bothrops asper (and other pit vipers) antivenom PoliVal-ICP from Costa Rica, and the B. lanceolatus-specific monovalent antivenom BothroFav (
Table 2). Our findings, sourced from a variety of in vitro assays, demonstrated that these three distinct products exhibit variable binding and neutralisation potencies against the seven venoms under study, though in the majority of experiments each antivenom showed clear evidence of venom toxin recognition and at least some inhibition of functional activities. Collectively our findings suggest that the PoliVap-ICP and BothroFav antivenoms show particular promise for wider therapeutic use in regions where the current provision of specific antivenom is limited. The data presented here strongly advocate for future studies to preclinically validate our findings of paraspecific efficacy to facilitate downstream clinical use of these antivenoms in snakebite victims. 3. DiscussionPit vipers of the genus Bothrops cause most of the severe snakebites that occur in the Caribbean, Central America and South America [
1,
3]. Snakebite envenoming by Bothrops species manifests as potentially lethal systemic haemorrhage and coagulopathy, while extensive local tissue damage and inflammation occurs frequently and can lead to amputation and other irreversible local sequelae resulting in life-long morbidity [
1,
3]. The WHO guidelines specify that snakebites should be treated with antivenom, the only venom-specific therapeutics that exist today. However, this course of action is difficult to implement in disadvantaged regions with low antivenom availability, accessibility and/or affordability. Further, venom variation means that commercial antivenoms have limited geographical utility, and as such there are large parts of the tropical world for which no bespoke antivenom is manufactured, meaning that assessing the efficacy of existing products against snake species or populations for which no specific treatment exists (i.e., assessing their paraspecific efficacy) is of utmost importance [
24,
27]. Consequently, in this study we explored the extent to which three commercially available Central/South American antivenoms might have broad utility against Bothrops envenomings in the region.Our study used seven different Bothrops venoms from five different species that covered a broad geographical distribution in Latin America and the Caribbean. In line with previous studies (summarised in [
11,
30]), the toxin compositions of these venom samples differed considerably, as observed by SDS-PAGE gel electrophoresis (
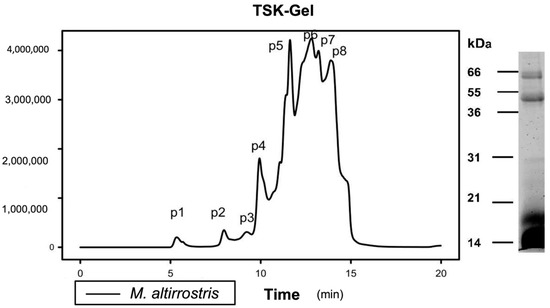
Figure 1A). However, all venoms exhibited clear presence of detectable proteins at three major molecular mass regions (7–12 kDa, 18–30 kDa and 55–60 kDa), and these observations may therefore reflect inter-specific differences in abundances and isoform numbers within the major viper toxin families, such as the SVMP, PLA2 and SVSPs, rather than major differences in the presence or absence of these different toxin types. Despite this compositional variation, some functional activities of the venoms appeared highly consistent in our study. For example, all seven venoms substantially reduced fibrinogen concentrations in venom-spiked platelet poor plasma (
Figure 5) and, while there were differences in potency, all were procoagulant to plasma (
Figure 4C). These findings are largely consistent with previous reports on the coagulopathic activity of the Bothrops venoms under study [
30,
31,
32], although Bourke et al. [
32] additionally showed via thromboelastometric approaches that the fibrinogenolytic potency of Bothrops venoms can vary considerably. We did observe variation in SVMP venom activity, as measured by enzymatic assay, with the Suriname and Colombian populations of B. atrox, along with B. jararaca venom, exhibiting considerably higher activity than the remaining venoms (
Figure 4A). These venom potency differences were not predicted based on the SDS-PAGE profiles and band intensity in the 55–60 kDa region (indicative of P-III SVMPs), or based on the literature, where B. lanceolatus, for example, has been reported to have the highest abundance of SVMPs in its venom (~75% [
12]), but exhibited a more modest enzymatic activity that the three venoms mentioned above.Despite these compositional and functional venom variations, the three anti-Bothrops antivenoms (Soro antibotrópico-crotálico, PoliVap-ICP and BothroFav) displayed extensive immunological recognition of these diverse toxin components, as clearly evidenced by both the Western blotting and ELISA experiments (
Figure 1,
Figure 2 and
Figure 3). The Western blots were non-discriminatory, perhaps surprisingly revealing that the monovalent BothroFav antivenom recognised a comparable diversity of toxins found across the various Bothrops venoms to the trivalent (though B. asper-specific in terms of Bothrops) PoliVap-ICP and the polyvalent Soro antibotrópico-crotálico antivenoms (
Figure 1). Endpoint titration and avidity ELISAs revealed some quantitative differences in antivenom-venom binding, with BothroFav generally exhibiting highest binding against the Caribbean and Suriname venoms from B. lanceolatus, B. caribbaeus and B. atrox, while PoliVap-ICP was superior against Central American B. asper and Colombian B. atrox venoms (
Figure 2 and
Figure 3). Interestingly, for each of these antivenoms we found evidence of paraspecific venom binding at comparable levels to that observed with the venom used for immunisation. While the Soro antibotrópico-crotálico antivenom was generally outperformed in these assays, most notably in the avidity ELISAs except against B. jararaca and B. atrox Suriname venoms, differences in immunological binding observed were relatively modest suggesting only a slightly reduced potency. Overall, these data provide strong support that each of these antivenoms contains antibodies capable of recognising and binding to a diverse range of toxins found in Latin American Bothrops venoms, irrespective of which venoms are used for the immunisation process.Because levels of antibody-toxin binding are not necessarily reflective of toxin inhibition, we used three different in vitro functional assays to explore paraspecific venom neutralisation by the three antivenoms. The fluorogenic SVMP activity assay revealed data highly consistent with the immunological assays, with all three antivenoms performing in a largely comparable manner, though Soro antibotrópico-crotálico exhibited the highest mean percentage reductions in venom SVMP activity (
Figure 4). Measures of venom-induced coagulopathy revealed a different inhibitory pattern, with Bothrofav and PoliVap-ICP exhibiting highly comparable neutralising potencies against procoagulant venom activities observed in the plasma assay, except that BothroFav was less effective against Colombian B. atrox and B. asper venom at the tested antivenom doses (
Figure 4). Measures of fibrinogen concentrations in venom-spiked PPP revealed a similar pattern, with Bothrofav and PoliVap-ICP exhibiting highly comparable inhibitory profiles, and in this case including equipotency against both B. atrox and B. asper (
Figure 5). Both antivenoms consistently outperformed Soro antibotrópico-crotálico in these two assays, suggesting that, despite extensive inhibition of enzymatic SVMP activity by this antivenom, perhaps other toxin types are contributing to the procoagulant venom phenotypes observed.This work highlights the therapeutic potential of BothroFav, PoliVap-ICP and, perhaps to a lesser extent, Soro antibotrópico-crotálico antivenoms for broad use against the venom of medically important Bothrops species found across Central and South America and the Caribbean. Most notably, across the various immunological and functional in vitro assays employed herein, we found only minor differences in the binding and inhibitory capabilities of these three antivenoms, despite distinct immunogens being used to raise the polyclonal antibodies (
Table 2). Each antivenom exhibited at least some capability to recognise, bind and inhibit the toxin activities of Bothrops snake venoms not used as immunogens, highlighting the potential paraspecific efficacy of such antivenoms, as also proposed elsewhere [
31,
33,
34,
35]. Overall, PoliVap-ICP performed well against most venoms tested in most assays, though perhaps surprisingly the monovalent product Bothrofav was highly comparable, apart from reductions in both binding and procoagulant toxin inhibition against Colombian B. atrox and B. asper venom. Except for the SVMP assay and for comparisons with the venom from B. jararaca used as an immunogen, the Soro antibotrópico-crotálico antivenom was generally outperformed by the other two products, though evidence of broad paraspecific cross-reactivity and toxin inhibition was observed. It is worth noting that in this study we were limited by using seven venoms sourced from five Bothrops species for our assessments of antivenom cross-reactivity. While these venoms span a broad geographical range (
Table 1), there are many other Bothrops species (the genus contains >50 species), including medically important species such as B. alternatus, B. moojeni and B. neuwiedi, and since venom variation can be unpredictable it remains unclear whether patterns of broad paraspecific binding observed here would extend to other members of this speciose genus [
24,
25,
36].Although promising, these findings must next be validated using more robust models of antivenom efficacy, including small animal models of envenoming, to assess whether these antivenoms are capable of significantly reducing the lethal, coagulant, haemorrhagic and dermonecrotic activities of various Bothrops snake venoms in vivo [
33,
34,
35,
37]. These analyses could perhaps also be usefully complemented by proteomic-based analyses of antibody binding (i.e., ‘antivenomics’), to determine which toxins are depleted by antivenoms and which are not recognised due to venom variation [
8,
38,
39]. Evidence of paraspecific preclinical efficacy and toxin depletion would provide a compelling basis to then explore the clinical efficacy of these antivenoms in observational studies of snakebite victims, particularly in regions of Latin America where species-specific antivenom therapies are currently not available. Expanding the clinical utility of existing antivenoms offers a potential short-term solution to reduce the severity of life-threatening systemic and morbidity-causing local envenoming caused by Bothrops pit vipers.


留言 (0)