1. IntroductionTetrodotoxin (TTX) is an extremely potent low-molecular-weight sodium channel blocker that originates from bacteria and is widespread in aquatic and terrestrial ecosystems. It is responsible for seafood poisoning events in the countries of the Indo-Pacific region [
1], where it was recorded to cause paralysis and respiratory and/or heart failure, in severe cases. Recently, invasive TTX-bearing species have become increasingly widely distributed in waters of Europe, North and South America, and Oceania, thereby expanding the geography of TTX poisoning [
2]. It has also been reported that in animals, TTX usually co-occurs with its analogues [
3], of which several are more potent than TTX itself [
4].Nemertea is a phylum of marine worms, also known as ribbon worms, comprising more than 1350 species [
5], of which most are active predators. Among nemerteans, TTX was first found in 1988 in Lineus fuscoviridis and Tubulanus punctatus [
6]. Subsequently, a number of TTX-bearing nemerteans were identified from all three classes of Nemertean (Palaeonemertea, Pilidiophora, and Hoplonemertea); those included extremely toxic species whose TTXs level reaches those recorded from pufferfish, octopus, and newts [
7,
8,
9,
10]. Currently, TTX profiles of different organisms attract the increasing attention of researchers aiming to elucidate the accumulation mechanisms and migration pathways of TTXs in ecosystems. Previously, a study of concentrations of TTX and its analogues in extremely toxic C. cf. simula specimens from different habitats revealed a wide variation in levels of toxins. Thus, using a mouse bioassay, which is a less precise method than high performance liquid chromatography with tandem mass spectrometry (HPLC–MS/MS), Asakawa et al. [
10] estimated the toxicity of the C. simula population that inhabits Hiroshima Bay (Japan) to be equivalent to 30.08–4555.02 µg/g of TTX per 1 g of body weight. Nevertheless, they did not provide information about concentrations of the TTX analogues separately. Another study of C. simula collected at Godrevy Point, Cornwall, England, described the proportions of separate TTX analogues without evaluating the range of TTXs concentrations, since TTXs were quantified in only a single specimen [
11]. An earlier published study with TTX quantification in a pooled sample from seven specimens of K. alborostrata, showed that it contained
12]. In another study, only qualitative TTXs detection was carried out.
In the present report, the concentrations of TTX and its analogues in two species of nemerteans, the only TTX-containing animals known from the Russian coast of the Sea of Japan were assessed. TTXs profiles for 32 specimens of C. cf. simula and 36 specimens of K. alborostrata, collected from Spokoynaya Bay, Peter the Great Bay, were analyzed, and based on the results, the possible way of their toxification was suggested.
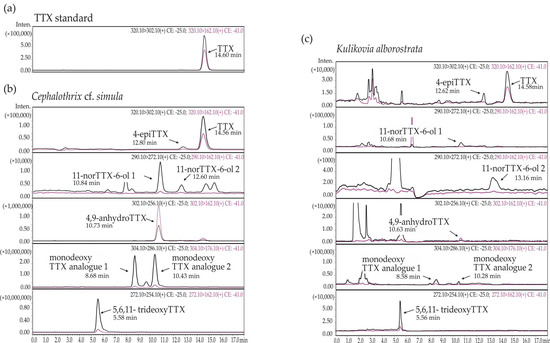
3. DiscussionAccording to the results of HPLC–MS/MS, the total concentration of TTXs in C. cf. simula, having a mean value of 1685.17 ± 1938.80 µg/g of nemertean body weight, varies by several orders of magnitude, from 85.75 to 7108.26 µg/g (
Table 1), which fits within the range of toxin concentrations reported for this species [
10,
11]. The TTX concentration in the extracts of K. alborostrata is 1.96 ± 2.18 ng/g (with a range from 0.35 to 8.11 ng/g), i.e., significantly lower than that in the extracts of C. cf. simula (
Table 1 and
Table 2). Therefore, the level of toxins in both C. cf. simula and K. alborostrata, within and between separate populations, can vary by several orders of magnitude. The high difference in the toxin concentration within the same species is typical for most TTX-bearing animals, like pufferfish, mollusks, newts, etc. [
13,
14,
15,
16,
17,
18,
19].The toxins TTX, monodeoxy TTX analogue 1, and 5,6,11-trideoxy-TTX in the C. cf. simula individuals, analyzed in the present study, have the highest percentages, from 11.78 to 78.08% for TTX (from 27.11 to 2677.51 µg/g), from 0.25 to 56.85% for monodeoxy TTX analogue 1 (3.03 to 1340.96 µg/g), and from 14.74 to 80.73% for 5,6,11-trideoxy-TTX (from 13.35 to 5737.72 µg/g). Despite the wide variation in toxin concentrations between individuals, the total proportion of the three toxins in all specimens made up 96.78 ± 3.09% of the total amount of TTXs. The percentages of TTX, monodeoxy TTX analogue 1, and 5,6,11-trideoxy-TTX in the extracts of seven C. cf. simula specimens, collected from the same locality (Peter the Great Bay, Sea of Japan) in previous years, were similar [
12,
20]. In the study of 2018, the TTX percentage amounted to 21.85%; monodeoxy TTX analogue 1, 32.33%; and 5,6,11-trideoxy-TTX, 39.93% (with the total proportion of 94.11%) [
12]. In 2020, the TTX percentage amounted to 33.63 ± 15.48%; monodeoxy TTX analogue 1, 17.37 ± 15.86%; and 5,6,11-trideoxy-TTX, 44.97 ± 14.72% (with the total of 97.36 ± 1.75%) [
20]. A different pattern of TTX analogues was recorded from a C. simula caught in England: the content of TTX was 64%, followed by 6,11-dideoxyTTX (21%), the total proportion of 5-deoxyTTX (11-deoxyTTX) and 5,6,11-trideoxyTTX was 9.9% [
11], and 11-oxoTTX amounted to 5%. Since only one specimen was investigated, it was impossible to draw any conclusions regarding toxin profiles of different C. cf. simula populations. Nevertheless, several studies carried out on pufferfish Lagocephalus sceleratus, collected off North Lebanon [
21] and along the Greece coast [
22], as well as Arothron nigropunctatus from two different localities, Okinawa, Japan, and the Solomon Islands [
23] have shown that neither qualitative nor quantitative TTX compositions can be considered stable characteristics of the species. TTXs have an exogenous origin in TTX-bearing animals, that accumulate directly from marine bacteria, the primary TTX producers, and/or through the food web [
24,
25]. Since no biotransformation of TTX and its non-equilibrium analogues has been observed in living organisms [
26], the intraspecies difference in qualitative and quantitative TTX profile, between specimens from different localities, may indicate its correlation with the source of toxins, which is supposed to be unique in each region. The characteristic toxin profile of each locality can originate from spectra of free-living bacteria and the microbiome of animals inhabiting it. The final TTX profile of consumers, including nemerteans, is presumably determined by their microbiome, or diet preferences (the TTX profiles of their prey items), or both. To date, there have been no studies considering the correlation between microbiome and TTX profile. However, the observations on the microbiomes of C. simula from England and C. cf. simula from the Sea of Japan have revealed differences in the most representative bacteria, even on the phylum level.An interspecies comparison of the TTX profiles of C. cf. simula and K. alborostrata have shown that some of the major toxins were the same. All the nemertean individuals were divided into six groups, based on the toxins which comprised 60% of all toxins recorded from them: (1) TTX; (2) 5,6,11-trideoxyTTX; (3) TTX + 5,6,11-trideoxyTTX; (4) TTX + monodeoxy TTX analogue 1; (5) 5,6,11-trideoxyTTX + monodeoxy TTX analogue 1; and (6) 5,6,11-trideoxyTTX + 11-norTTX-6-ol 2. Four groups were common for C. cf. simula and K. alborostrata, while both species were represented by five groups each (
Figure 3). The comparable compositions of the major toxins may result from similar accumulation pathways of TTX and its analogues for the two species, including obtainment from their own microbiomes and/or preferred diet. Recently, it has been reported that the microbiomes of C. cf. simula and K. alborostrata differ significantly [
27] and, although the role of microbiome cannot be ruled out, it is probable that the levels of TTXs within both species may also include contributions from consumed prey. The effect of prey toxins profiles on predator ones has already been demonstrated by several researchers. Thus, Ito et al. [
28] have shown that the TTX / 5,6,11-trideoxyTTX ratio in pufferfish (Chelonodon patoca) and toxic goby (Yongeichthys criniger) can result from this ratio in their presumable prey, the flatworm Planocera multitentaculata. In another study on Octopus vulgaris fed shellfish, containing another group of guanidinium toxins, paralytic shellfish toxins (PSTs), the similar PSTs prevailed as a result [
29]. Since in most individuals of C. cf. simula and K. alborostrata from the same locality the major toxins are similar, TTX and 5,6,11-trideoxyTTX (
Table 1 and
Table 2), which presumably suggests that their toxification have common sources. This suggestion is supported by data from articles describing similar nemerteans’ dietary preferences as predators. Thus, members of the family Lineidae (which includes K. alborostrata) prefer mostly polychaetes, from several families (Nereidae, Phyllodocidae, Polynoidae, and Terebellidae) [
30,
31,
32,
33,
34,
35]. Prey preferences of the Cephalothrix species are poorly known, but several feeding experiments have revealed their diverse diet that includes a wide range of taxonomic classes of prey: polychaetes [
36], oligochaetes, nematodes [
33], and crustaceans (amphipods and isopods) [
37]. Therefore, it can be assumed that several prey items, common for both nemertean species, were the sources of the same toxins in them. Nevertheless, this assumption should be further verified through dietary investigations, using DNA metabarcoding and determination of toxin profiles of the presumed prey. Additionally, the contribution of the microbiomes of C. cf. simula and K. alborostrata to their toxification should not be ignored, and remains an important issue to address.


留言 (0)