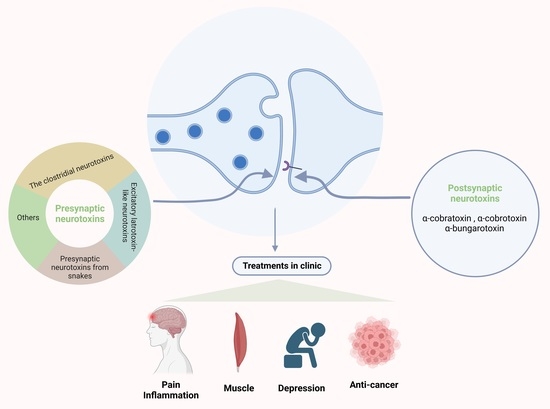
The postsynaptic neurotoxins have postsynaptic blocking functions at the skeletal muscle end-plate and neurons, named α-neurotoxins [
15]. They act as antagonists of nAChRs in the NMJs and neurons [
109]. The α-neurotoxin family has a similar mechanism of the function that causes muscular paralysis due to blockade of the postsynaptic nAChRs at the NMJ [
110,
111,
112].The α-neurotoxins in snake venoms belong to the three-finger toxins [
113]. These kinds of toxins are flat molecules, and the reason for the name of “three-finger” is that the flat molecules have a small globular hydrophobic core, and this core can form three adjacent loops, with the shape of three fingers [
111,
114,
115]. The three-finger toxins are mainly generated form Elapid snakes, including α-cobratoxin (α-Cbtx), α-cobrotoxin (α-Cotx), and α-bungarotoxin (α-Bgtx) [
116]. In addition, based on their molecular structures, α-neurotoxins are roughly divided into three categories: short-chain toxins with 60–62 amino acids residues and four disulphide bridges (α-Cotx) [
117]; long-chain toxins with 66–75 amino acid residues and five disulphide bridges (α-Cbtx) [
118]; and weak toxins, which have the feature of hypotoxicity (LD50 ~ 5–80 mg/kg as compared to LD50 ~ 0.04–0.3 mg/kg for other toxins) [
103]. What they have in common is that they all target muscle α1 nAChRs, but only long-chain α-neurotoxins can bind to the α7 nAChR with high affinity [
109,
119,
120], while only the dimmer of cobrotoxin binds to it. As we know, the α7-type nAChR is considered as a potentially essential target for the treatment of many diseases in the clinic [
121], such as cognitive disorders [
122,
123], inflammatory diseases, and chronic pain [
124]. Alzheimer‘s disease, Parkinson’s disease, Lewy body dementia, and schizophrenia all have the feature of neuroinflammation [
125,
126,
127,
128,
129,
130,
131,
132,
133]. At present, some researchers have started to explore the effect of α-neurotoxins on central nervous system diseases [
134,
135,
136,
137,
138]. Among these toxins, the α-bungarotoxin (αBgt) is the most popular one as a pharmacological tool used for studying α7 nAChR [
121] because of the practically irreversible binding to the Torpedo acetylcholine receptors [
139].In addition to snake neurotoxins, α-conotoxins generated from marine organisms of the genus Colocasia also target nAChRs [
140]. There are many subgroups of α-conotoxins that are selective for different subtypes of AChRs, such as α3/5-conotoxins that selectively block the muscle nAChR, and α4/3-, α4/4-, α4/5-, α4/6-, and α4/7-conotoxins that selectively block the neuronal nAChRs [
140,
141,
142] (
Figure 2).
All the above-mentioned neurotoxins that are classified into presynaptic and postsynaptic neurotoxins, even though they are from different sources, have many types, and their complex subtypes and structures make it difficult to generalize with a symbolic model that they have a similar principle of function in the intervention of the neuromuscular signal transmission process, in which they affect the transmission of neurotransmitters at synaptic sites by regulating ions, receptors, and membrane fusion proteins and in other ways.
Besides those mentioned above, there are still some toxins not marked as “neurotoxins” that exert the same action on the nervous system. For example, the epsilon toxin (ET), produced by clostridium perfringens types B and D, can cause enterotoxaemia in sheep, goats, and cattle. When it enters the brain, it targets the neural cells and also induces the release of glutamate and other transmitters; but the specific mechanism needs more exploration [
143]. Some bacterial enterotoxins, such as toxin B (TcdB) and cholera toxin (CT), can attack the enteric nervous/endocrine system. There is evidence that CT activates a secretomotor neural reflex to further enhance the secretory responses; and TcdB can catalyze the glucosylation of Rho proteins (Rho, Rac, and Cdc42) to block neurotransmitter exocytosis [
30,
144,
145,
146,
147].


留言 (0)