1. IntroductionThe highly pathogenic avian influenza virus (HPAIv) has caused worldwide epidemics in poultry and humans [
1]. The H5N1 influenza virus first affected humans in 1997; in 2021, it spread at an unprecedented rate throughout the east coasts of Canada and the United States. Since late 2003, H5N1 has become the most contagious and deadly pathogen in domestic fowl and wild birds, as well as in human populations in Asia, the Middle East, Eastern Europe, and Africa. Over 800 human cases have been recorded, with a fatality rate of more than 50% [
1,
2]. Hemagglutinin (HA) is a major envelope glycoprotein on the surface of the influenza virus. By mediating the fusion of the endosomal membrane through interaction with the sialic acid receptors on target cells, the HA trimer initiates the viral infection [
3,
4,
5,
6]. Therefore, HA proteins have been utilized as primary targets for the influenza vaccine and antibody development to neutralize the influenza virus. Therapeutic approaches that interfere with the HA protein on the influenza virus have demonstrated excellent anti-viral activity in clinical trials [
3,
7,
8].Camelids as domesticated animals are significant for the economy in many regions of the world and can adapt to a wide range of extreme ecosystems [
9]. Recent research on camelid B cells has shown that all individuals of this species possess a special immunological trait that is uncommon in other species [
10]. A ‘nanobody’ is a novel single-domain antibody derived from the variable domain of the heavy chain, from heavy-chain-only antibodies (VHH) in camelids. Due to the unique biochemical characteristics, including small size, high affinity, low cytotoxicity, as well as deep tissue penetration [
11], nanobodies have been considered the “next generation” antibodies [
12]. Recently, there has been an increased interest in camelid immunology. Considerable research has been devoted to studying immunoglobulin and nanobody development [
13,
14,
15]. However, the immune system consists of a complex network of immune cells and related cytokines that contribute to efficient immune responses against infections. Studies on the immune response to antigen-specific immunization are important to understand the immune system of camelid animals.To date, only a few studies have investigated the cellular components of the camelid immune system. Several studies have described the diversity of peripheral blood mononuclear cells (PBMCs) and revealed the relative proportions of the primary leukocyte subpopulations in camelid animals using flow cytometry [
9,
16]. Due to the small number of identified cell subgroups and the inconsistent phenotypic and functional definition of leukocyte composition, it is challenging to accurately compare the immunophenotype of leukocytes obtained in different studies [
17,
18,
19]. Single-cell RNA sequencing (scRNA-seq) technology offers unprecedented precision for describing novel cell types, cell states varying from healthy to pathological, or immune cell responses to antigen stimulation [
20]. Sophisticated scRNA-seq technology has been used to analyze the kinetics of the immune response during pathogen infections and to reveal the immune regulation mechanism at the single-cell level [
21].
In this study, we collected PBMCs from an alpaca and described the transcriptome landscape and longitudinal alterations in response to H1N1-HA protein immunization at the single-cell level. A total of 35,853 cells were obtained during the pre- and post-immunization stages. Based on transcriptome data, we defined the immunophenotype of leukocytes and discovered that HA antigen boosting triggered both innate and adaptive immune responses in the alpaca. Characterization of important lymphocyte subpopulations and dynamics of the immune response to antigen-specific immunization offer valuable information for the development of potent nanobodies and influenza vaccines in camelid animals.
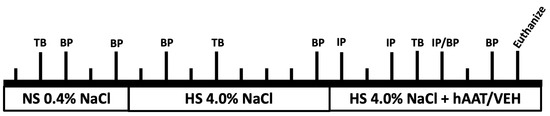
2. Materials and Methods 2.1. Alpaca ImmunizationA young, healthy adult female alpaca raised in the China National Gene Bank was immunized twice over a period of 14 days. For immunization, the animal was injected with 250 µg of influenza A hemagglutinin protein (HA1-V5229-1mg, ACROBiosystems, Newark, DE, USA) mixed 1:1 with complete/incomplete Freund’s adjuvant. Whole blood samples were collected before immunization (day 0, HA-0), and at day 17 (HA-1) and day 19 (HA-2) after the second immunization (
Figure 1A). 2.2. Serum Titer Assay
An HA-specific enzyme-linked immunosorbent assay (ELISA) binding assay was performed to determine the antibody titer in the alpaca serum. Maxisorp ELISA plates (Invitrogen) were coated with 100 ng of HA protein in a coating buffer (100 nM NaHCO3 in phosphate-buffered saline (PBS), pH 8.3) and incubated overnight at 4 °C, with corresponding blank (non-antigen containing) control and negative (irrelevant antigen containing) control sets. Next, 200 μL of blocking buffer (2% skimmed milk in PBS) was added for 1 h at 25 °C. The alpaca serum collected at HA-0, HA-1, and HA-2 was serially diluted from 101- to 108-fold with phosphate-buffered saline (PBS) and incubated in the plate at 25 °C for 1 h. Next, the plates were washed five times with PBST (0.05% Tween-20 in PBS) before adding the secondary antibody. HRP-conjugated mouse anti-alpaca IgG-antibody (Abcam) was added at a 1:2000 dilution and incubated for 1 h at 25 °C. After adding 100 μL TMB substrate (Abcam, Cambridge, UK), the absorbance at 450 nm was read within 15 min using Synergy™ H1 (BioTek, Winowski, VT, USA).
2.3. Peripheral Blood Mononuclear Cell (PBMC) Isolation
Whole blood samples were collected, and peripheral blood mononuclear cells (PBMCs) from each sample were isolated using Ficoll-Paque™ PLUS Media (GE Healthcare, Chicago, IL, USA) within 2 h, according to the manufacturer’s instructions. Briefly, 10 mL of whole blood was transferred from the collection vessel to a 50 mL EP tube; an equal volume of PBS solution was added to the EP tube to dilute the blood. Then, the diluted blood was added to 15 mL of Ficoll and centrifuged at 500× g for 20 min (accelerate 3, decelerate 0); next, the buffy coat was carefully transferred to a new tube and diluted with PBS to a total volume of 20 mL. The samples were centrifuged again at 600× g for 6 min, after which the buffer was discarded, and the cell pellet was resuspended in 3 mL PBS.
2.4. Single-Cell Library Construction and Sequencing
The DNBelab C Kit (MGI, #1000021082) was used to construct the library. Isolated PBMCs were resuspended as single cells at a density of 5000 cells/mL. Cells were wrapped in droplets with a negative pressure chip, and mRNA was transcribed to cDNA to generate a sequencing library according to the manufacturer’s instructions. Sequencing libraries were quantified using a Qubit™ ssDNA Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA). The cDNA libraries were then subjected to DIPSEQ T1 sequencing (MGI).
2.5. scRNA-Seq Data ProcessingHigh-quality scRNA-seq data with valid barcodes were aligned to the genome of VicPac3.1 through the STAR software, and a unique molecular identifier (UMI) count matrix was generated using PISA (version 1.10.2) (
https://github.com/shiquan/PISA (accessed on 8 August 2021)). The raw transcript count matrix was loaded into the R (v4.0) software using the Seurat (v3.1.5) package [
22]. 2.6. Data Integrating and Cell ClusteringWe used the R package Seurat 3.1.5 to integrate and analyze datasets from the three samples (HA-0, HA-1, and HA-2). The integrated mRNA expression matrix was first filtered following the Seurat recommendation and a total of 35,853 cells with unique UMI were obtained [
23]. Unsupervised clustering was conducted using Seurat with the parameter res = 0.5, which revealed a total of 18 clusters. We used mRNA biomarkers obtained from recently published articles to classify these clusters into seven major groups (
Supplemental Table S1) [
24]. 2.7. Differentially Expressed Gene (DEG) Analysis
DEGs were calculated using the function FindMarkers built in Seurat with the default parameters. The resulting DEGs were filtered with p_val_adj < 0.05 and then sorted according to the average log2 fold change (avg_log2FC).
2.8. Gene Ontology AnalysisWe used the clusterProfiler to annotate the functions of the cell subsets. We filtered the enriched pathways with an FDR q-val of ≤0.05 [
25]. The pathways that normalized the enrichment score in the top 20 are shown in the results. 4. DiscussionStudies on camelid immunity have mainly focused on the generation of nanobodies, which have been widely used in therapeutics and diagnostics [
64,
65]. Few studies have examined the characteristics of alpaca immune cell composition in response to antigen immunization. In this study, we constructed a comprehensive single-cell landscape of peripheral immune cells from an alpaca with HA antigen stimulation. Using single-cell RNA sequencing, we profiled 35,853 immune cells sampled before and after immunizations. Immune cells were classified into seven major clusters, including CD4+ T cells, naive T cells, monocytes, natural killer cells, and dendritic cells. The immune responses of PBMC clusters were analyzed in detail separately. It is crucial to systematically identify the characteristics of immune cells in response to antigen-specific immunization, and this dataset will undoubtedly further elucidate the underlying molecular mechanisms of the unique immune system of camelid animals.Antigen-mediated induction of antigen-specific B and T-cell responses requires the activation of the innate immune system, particularly with respect to antigen-presenting cells [
66]. Following HA immunization, we found that the proportion of CD14+ monocytes steadily increased to >50%. Two subsets of CD14+ monocytes were activated and displayed pro-inflammatory characteristics by upregulating the expression of genes related to cell activation and differentiation, cell trafficking, and cytokine production. DEG analysis revealed that monocytes were mostly enriched in activities of antigen presentation and granulocyte chemotaxis pathways, which indicates that the innate and adaptive immune cells were activated in response to HA stimulation in the alpaca. However, the activation of the innate immune system can be both protective and detrimental during infections [
67]. HPAIv infections can induce a cytokine storm and exaggerate innate immune response, which results in severe pneumonia or death [
26,
62,
68,
69]. The substantial increase in monocyte percentage and cytokine production seen in response to HA immunization may indicate an excessive infiltration of pro-inflammatory monocytes, which may result in immunopathology [
26,
70]. This discovery offers a different perspective to illustrate the generation of cytokine storms.In addition to the monocytes, the proportion of CD8+ activated T cells also increased. GO analysis revealed that the T cell activation pathway and expression levels of cytotoxicity-related genes were upregulated in CD8+ activated T cells. These results demonstrated that HA-immunization activated the innate immune system, which could assist T cell activation against the infection. We have also noticed that part of the CCR7+SELL+ CD4+ naive T cell cluster identified in this study simultaneously expressed T cell activating genes such as CTLA4 and LTB. This phenomenon emphasizes that naive T cells are much more heterogeneous than previously understood [
71,
72,
73,
74,
75]. These CCR7+CTLA4+ T cells were evenly distributed among the HA-0, HA-1, and HA-2 samples (data not shown), indicating that the expression of CTLA4 CD4+ in naive T cells was not induced by HA immunization. We speculate that the co-expression of CCR7 and CTLA4 in these CD4+ naive T cells might be a result of continuous stimulation by other antigens several months prior to HA immunization. Further studies on the immune landscape of camelid animals are needed to reveal the occurrence of these naive T cells.Four B cell subclusters, including naïve B cells, CD19low B cells, SELL− B cells, and plasma B cells were identified in the alpaca PBMCs. This study revealed that SELL− B cells highly express the FCRL4 gene, which may inhibit memory B cells from differentiating into plasma cells by reducing the proliferation and differentiation potential [
76]. SELL− B cells were assumed to have an atypical memory B cell phenotype following specific antigen stimulation [
77]. A high proportion of plasma B cells highly expressed antibody secretion-related genes, such as TNFRSF17 (BCMA), FKBP11, and MZB1, which can be utilized to develop antibodies targeting alpaca plasma B cells. This is helpful for alpaca plasma cell enrichment during nanobody development. The proportion of plasma B cells during antigen immunization determines the effectiveness of antigen-specific antibody generation [
78,
79,
80]. In this study, we found that the proportions of plasma B cells dramatically increased by day 17 but were significantly reduced by day 19 (day 5 after the second HA immunization), suggesting that the proportion of plasma B cells would reach a peak at three days after the second immunization. Plasma cell dynamics during the specific antigen immunization should offer valuable references for nanobody discovery in immunized camelids [
79,
80]. However, the sampling periods between the first and second immunizations were not studied here. A larger and more thorough cohort of camelid animals should be constructed in the future to accurately identify the molecular mechanisms underlying the immune response.

留言 (0)