
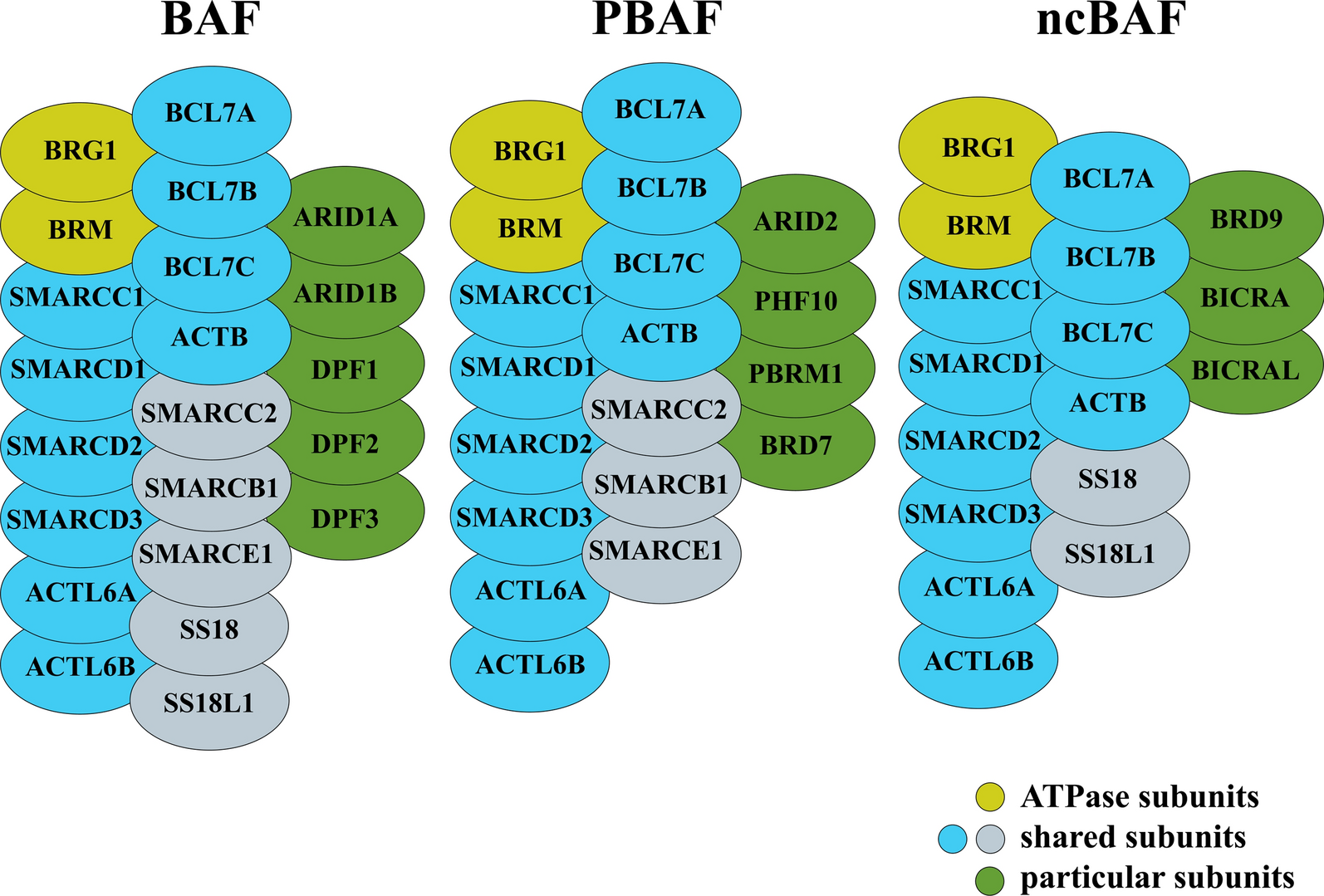
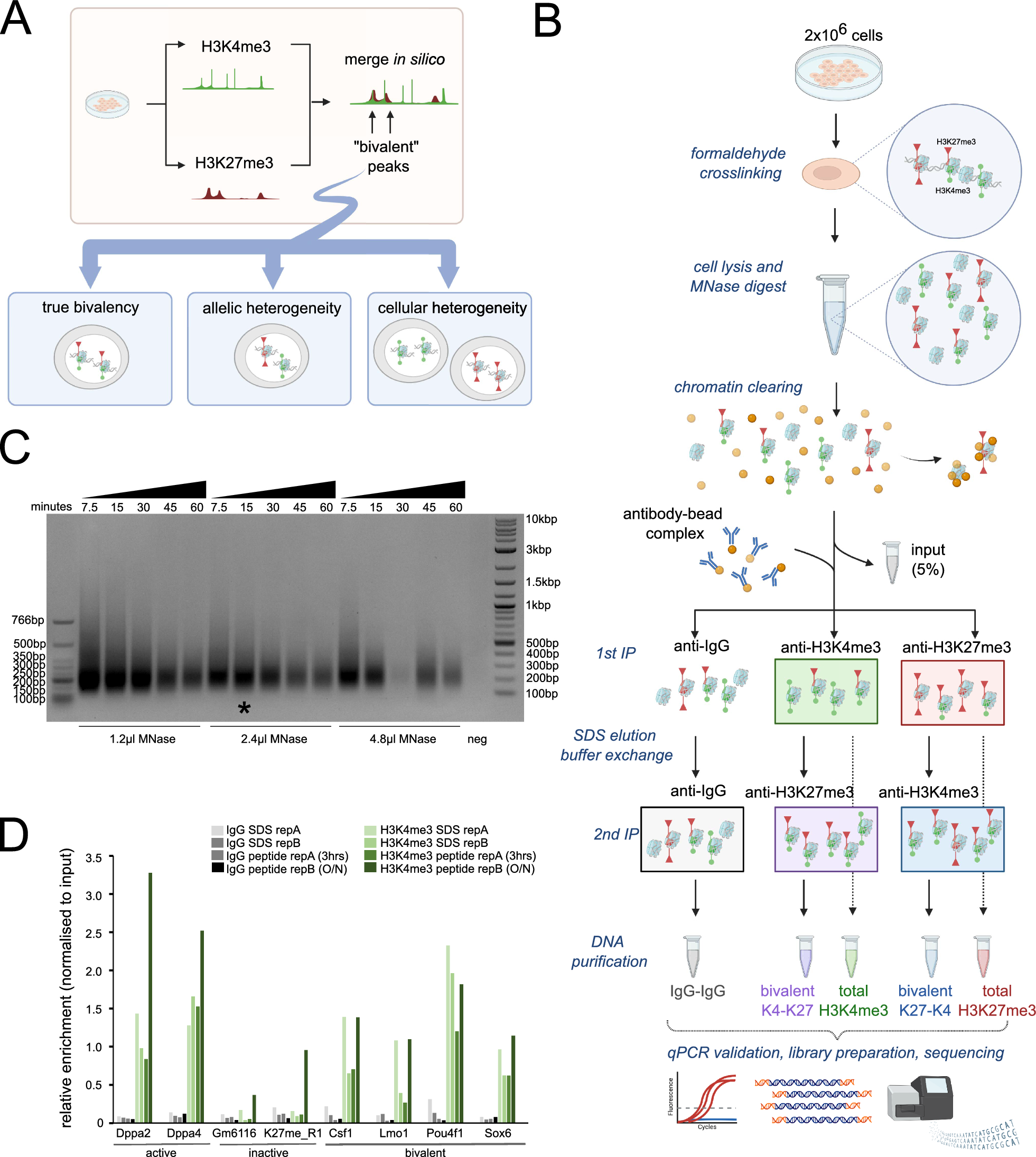
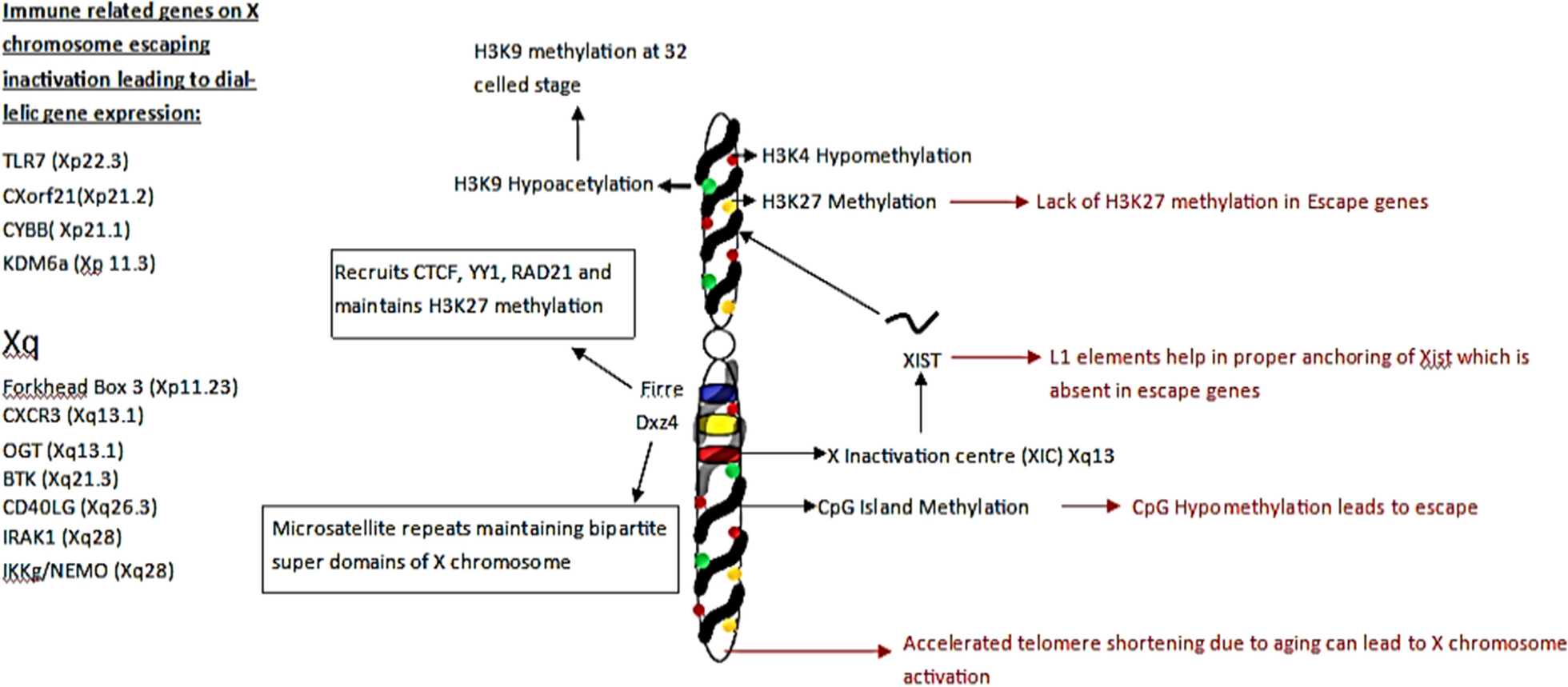
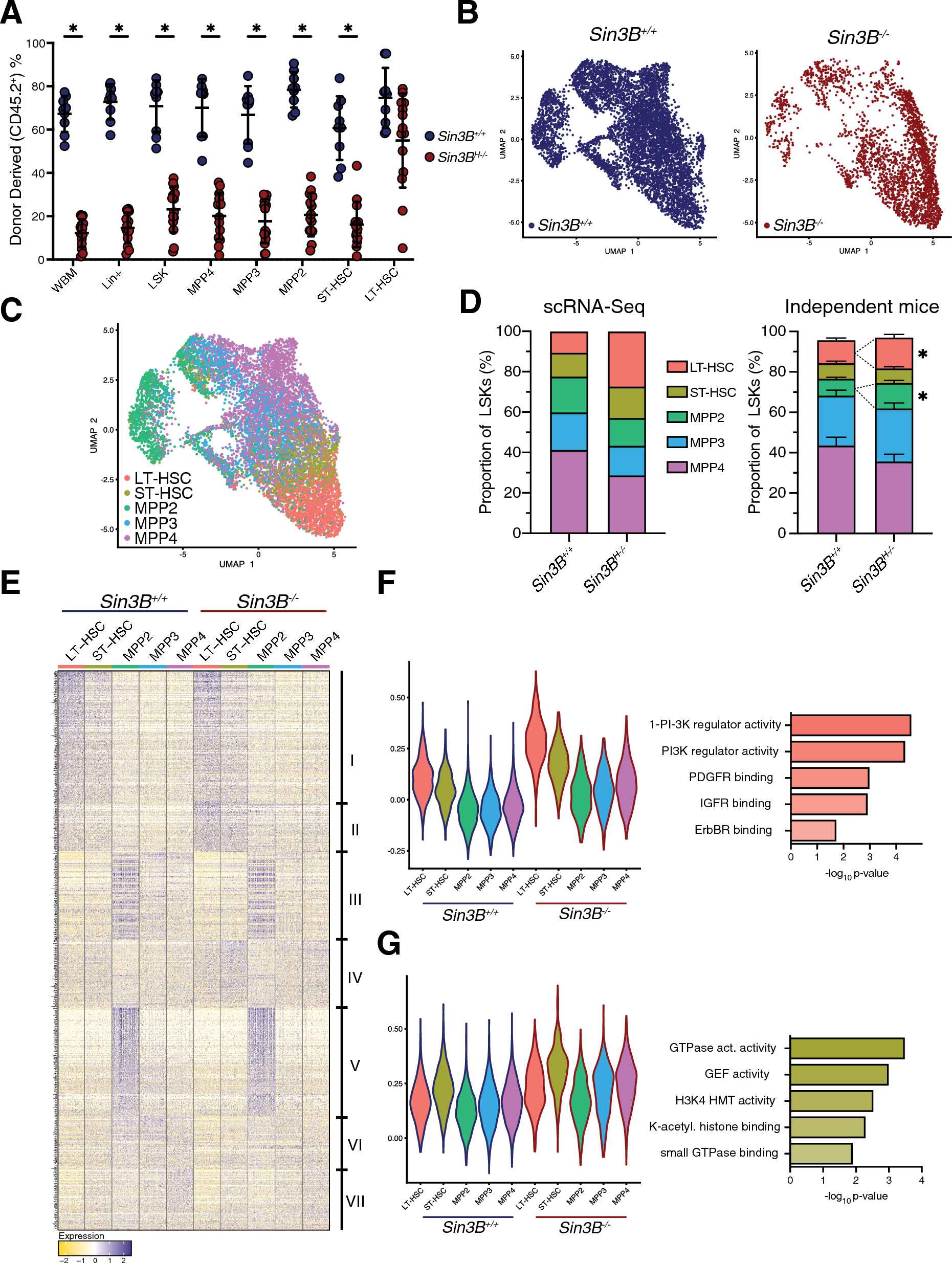
記住我


To obtain liver spheroids from HepG2/C3A cells, we optimized the protocol originally described by Wrzesinski and colleagues [11] for ease of use and to achieve reproducible and cost-effective culturing of > 100 spheroids per bioreactor. In brief, the culture was initiated by utilizing HepG2/C3A flat culture (2D culture) grown to 80% confluency (Fig. 1A). To culture the cells as spheroids (3D culture), 1.2 × 106 cells were plated in an ultra-low attachment plate containing microwells (approximately 2,000 cells per microwell) and were incubated for 24 h to allow them to self-aggregate (Fig. 1B). Before transfer to a bioreactor, the size and roundness of spheroids were analyzed by microscopy to ensure a uniform culture. Newly formed HepG2/C3A spheroids (Fig. 1C) ranged from 152 to 232 µm (Additional file 1: Fig. S1A), with an average size of 201 µm (± 18.3 µm). As spheroids get older, they get bigger and present a denser core compared to the borders (Fig. 1D and E). Although they are morphologically formed after 2 weeks, they only reach a functionally active plateau after 3 weeks in culture, as previously demonstrated [12]. The spheroid shown in Fig. 1F, which was maintained in culture for 22 days, represents a functionally mature spheroid.
Fig. 1
The proteome and metabolome of HepG2/C3A spheroids resemble a functional liver. A HepG2/C3A (human hepatocellular carcinoma) flat cells were cultivated in DMEM media supplemented with 10% FBS, until colonies reached 80% confluence. B Then, cells were trypsinized and plated on an ultra-low attachment 24-well plate containing microwells. C Spheroids were then detached from the plate, transferred to a bioreactor, and cultivated in the same media as flat cells (day 0). D Spheroids on day 5, E day 13, and F day 22. Images were acquired using PAULA Smart Cell Imager (Leica) with 10 × magnification. Scale bar = 100 µm. G Clustered network of the 200 upregulated proteins in HepG2/C3A spheroids. Clusters reflect connected portions of the network and correspond to functional categories of the proteins. The size of nodes represents p-value, color darkness represents the score (fold change enrichment times the p-value), and line thickness represents the score of interaction confidence retrieved from the software String (v11, https://string-db.org). The network was constructed by Cytoscape [20]. H Bar graphs show the daily secretion levels of the bile salts taurodeoxycholate, tauro α-muricholate, and taurochenodeoxycholate detected in the supernatant of spheroid culture
By growing HepG2/C3A cells as 3D spheroids, we verified whether the expressed proteome resembled more closely the one of functional liver. Biological processes significantly enriched in HepG2/C3A spheroids revealed proteins mostly related to the carbohydrate, cholesterol, and vitamin B6 metabolism as well as proteins related to amino acid metabolism and detoxification of metal ions (Fig. 1G). In fact, besides their role in metal detoxification, metallothioneins also contribute to the antioxidant activity and protective effects against free radicals performed by the liver [18]. We identified 5 isoforms of metallothioneins (MT1E, MT1F, MT1G, MT1X, and MT2A), all more abundant in spheroids in comparison to flat cells (i.e., grown in 2D culture) (Additional file 1: Fig. S1B). We were also able to detect some biomarkers known to be predominant in the liver, whether in disease or normal conditions, such as alpha-fetoprotein (AFP), albumin (ALB), apolipoprotein E (APOE), lactate dehydrogenase (LDHA), alanine aminotransferase (GPT2), and aspartate aminotransferase (GOT2) (Additional file 1: Fig. S1C). It is worth pointing out that our data were log-transformed and normalized to correct for any artificial biases in sample pipetting and injection that consequently could lead to differences in protein content. Therefore, the differences seen in the number of proteins between flat cells and spheroids indicate relative changes of protein expression and not an overall higher content of protein in the spheroids.
We also aimed to demonstrate that the longer spheroids are cultured, the more they produce and secrete metabolites typical of liver functions. Metabolic profiling of collected cell media from spheroid cultures revealed increasing levels of the bile salts taurodeoxycholate, tauro α-muricholate, and taurochenodeoxycholate (Fig. 1H), which are known to be produced by the liver. The full metabolite profiling of the cell media is available as Additional file 3: Table S1.
Together, these results confirmed that HepG2/C3A cells grown using the ClinoStar (CelVivo) 3D technology resemble the typical characteristic of functional liver, which includes maintaining whole-body lipid, glucose and energy metabolism, as well as detoxification of drugs, xenobiotics, and metals [19].
Liver spheroids respond to chromatin decondensationTo evaluate the sensitivity of our 3D model to drug treatment, spheroids were maintained in a growth media containing 20 mM of sodium butyrate (NaBut) for 3 days (Fig. 2A). After the collection of spheroids and culture supernatant, the treatment media were replaced by the standard growth media, and spheroids were maintained in culture for additional 7 days, to evaluate whether they could recover their normal physiology. Sodium butyrate is known to act as a histone deacetylase inhibitor (HDACi), favoring histone acetylation and thus remodeling chromatin toward an open and transcriptionally active state [21]. HDACi can also have anti-cancer effects, which depend on the type of cancer and the treatment dose [22]. We analyzed histone modifications by mass spectrometry, using the protocol optimized by Sidoli and Garcia [23]. Obtained chromatograms were extracted using EpiProfile [24].
Fig. 2
Spheroid respond to chromatin decondensation. Spheroids were treated with 20 mM of NaBut and were kept in culture for 3 days. After collection of 5 spheroids per time point, the treatment media were replaced by the standard growth media and the remaining spheroids were maintained for additional 7 days, until the last collection on day 10. Histones were extracted from 5 spheroids and analyzed by mass spectrometry. A Workflow for spheroids treatment. B Total levels of histone peptides containing 1, 2, or 3 methylations (me1, me2, me3, respectively) or containing acetylations (ac). C Total levels of histone H4 peptides containing acetylations (1ac, 2ac, 3ac, 4ac). Unmod, unmodified peptide. * p < 0.05, ** p < 0.005, and *** p < 0.0005 when compared with the Nt group. D Relative abundance of histone H3 acetylated peptides. Data are represented as means ± SD. Nt, non-treated. E Volcano plot representing NaBut vs Nt fold change after 3 days of treatment and F Correlation coefficient (R) of histone marks between non-treated (Nt) spheroids and NaBut-treated spheroids after recovery (Day 10). Highlighted green dots correspond to acetylated histone peptides
Overall, the total levels of mono, di, or trimethylated histones (me1, me2, me3, respectively) did not differ between non-treated (Nt) and NaBut-treated groups (Fig. 2B and Additional file 1: Fig. S2A). However, the acetylation (ac) levels increased after treatment (Fig. 2B). By using our protocol, the raw data for histone H4 are processed so that one particular peptide, which can carry up to 4 acetyl groups (amino acids 4 to 17, modification sites being K5, K8, K12, and K16), is highlighted. Therefore, we focused on that peptide to evaluate the accumulation of hyperacetylation on histone proteins. Treatment with NaBut for 3 days increased the levels of histone H4 peptides containing multiple acetylations, i.e. 3ac and 4ac, by almost 20% (Fig. 2C). Hyperacetylation of histones, triggered by NaBut, occurs slower in spheroids compared to flat cells (Additional file 1: Fig. S2B), which is expected because the lower proliferation rate of cells in the 3D culture leads to a slower production of new histones to be modified. Acetylation marks on histone H3 peptides were also quantified, and a similar trend was identified (Fig. 2D and Additional file 1: Fig. S2C, respectively). H3K9ac, as well as the combinatorial PTMs H3K9acK14ac and H3K18acK23ac, were the marks that were most upregulated by the treatment.
Importantly, following cessation of Na But treatment, we demonstrated that spheroids were able to reestablish their pre-treatment levels of histone modification after 7 days in the normal growth media (i.e., with no NaBut). By determining cell viability (adenylate kinase release), we demonstrated that spheroids were able to recover from the treatment as shown by the reduced number of damaged cells (Additional file 1: Fig. S3A). Furthermore, it is evident that on day 10, the levels of histone marks are not significantly different from their levels in the non-treated (Nt) condition (Fig. 2B–D). The recovery event is shown in Fig. 2E and F; the volcano plot clearly demonstrates that spheroids treated for 3 days show an enrichment of histone H4 hyperacetylated peptides (Fig. 2E), while on day 10 the levels return to basal levels and show a high correlation (R2 = 0.9228) with the non-treated condition (Fig. 2F). Besides showing that spheroids recover their normal metabolism and revert the effects caused by NaBut, our results suggest that histone acetylation seems to have minimal memory in our cell culture, as it is not being conserved once treatment has been withdrawn.
It is worth mentioning that this analysis is not possible in 2D cell monolayer, as 10 days is a sufficient time frame to fully replicate all the cells in culture, i.e., the recovery analysis would be performed on daughter cells rather than those which received the treatment. This highlights the relevance of the 3D culture model described here. To demonstrate that cells in 3D spheroids have a very slow replication rate, we quantified the relative abundance of the enzyme DNMT1, which is known for its role in maintaining methylation pattern in newly synthesized DNA strand [25]. We demonstrated that the levels of DNMT1 are lower in spheroids compared to flat cells (Additional file 1: Fig. S3B). Moreover, by metabolic labeling we confirmed that the protein turnover rate is significantly higher in flat cells than in spheroids, i.e., approx. 20% labeling after 2 days vs 1% labeling of proteins in spheroids (Additional file 1: Fig. S3C). This demonstrates that cells in 3D spheroids do not produce new DNA and new proteins, phenomena commonly associated with cell proliferation.
Transcriptional alteration and recovery from chromatin decondensationWe had demonstrated that NaBut treatment induced histone hyperacetylation (Fig. 2B–F). Since histone acetylation is known to directly and indirectly induce transcriptional activation [26], we aimed to demonstrate whether gene expression is affected by our treatment and whether it is restored once the treatment is removed. We performed RNA sequencing (RNA-seq) of treated and non-treated spheroids, identifying 6,230 genes with significantly different (FDR < 0.05) trajectories due to NaBut treatment (Fig. 3A). K-means clustering of this set of genes broke them down into 8 clusters, where genes in clusters A–D are initially upregulated and genes in clusters E–G following the opposite pattern over the time-course (Additional file 4: Table S2). Plotting the data onto a heat map, this clearly showed distinct transcriptional profiles between Nt and NaBut-treated spheroids over the time-course (Fig. 3A), highlighting that the majority of genes were upregulated in the presence of chromatin hyperacetylation. Although the clusters show different trajectories after treatment, nearly all affected genes return to the control levels by day 10 (NaBut10) where the spheroids were allowed to recover for 7 days (Fig. 3B). In order to get an insight into the altered biological processes induced by histone hyperacetylation and chromatin decondensation, over-representation analysis for each of the clusters was performed using either the Gene Ontology or Molecular Signatures databases. Biological processes that were impaired by the treatment represented a slowing of cell division and metabolism, such as cholesterol metabolism (clusters G and H), DNA replication-related chromatin changes (clusters E and F), and alcohol/sterol metabolism (cluster C) (Fig. 3C). On the other hand, a myriad of seemingly unrelated biological processes were enhanced by the treatment which included macroautophagy (cluster A), cell–cell adhesion and chemotaxis (cluster B), calcium activity (cluster C), and phospholipid/glycerolipid biosynthesis (cluster H). This indicates that chromatin decondensation induced by non-specific hyperacetylation led to a spurious upregulation of genes rather than a coordinated response to the treatment. To confirm this, enrichment for cell–type signatures showed loss of liver cell identity after treatment, such as downregulation of the hepatocyte and hepatoblasts (clusters E–H) and the upregulation of non-liver signatures, such as macrophage (cluster A), neurons (clusters B–C) microglia, astrocytes, heart, and lung (clusters C–D) (Fig. 3D).
Fig. 3
Transcription alteration and recovery following NaBut treatment. A Heat map showing genes significantly altered over the time-course of the treatment. Non-treated (Nt1, Nt2, Nt3, Nt10) and NaBut-treated (NaBut1, NaBut2, NaBut3, NaBut10) spheroids, treated for 1, 2, or 3 days and the recovery time point (day 10); n = 3. B Average trajectory of genes over the time-course for each cluster showing return to baseline levels at recovery time point (day 10). Ribbons represents standard error. C Gene ontology biological process over-representation analysis for each of the clusters. D Cell-type signature over-representation analysis for each of the clusters. Clusters A–D are downregulated and clusters E–H are upregulated over the time-course
Histone hyperacetylation and chromatin decondensation have a direct effect on the spheroids’ proteomeGiven the effect of NaBut on cellular biological processes at the transcriptome level, we then investigated the gene ontology (GO) enrichment at the proteome level (Fig. 4). GO analysis of upregulated proteins of spheroids treated with NaBut for 1, 2, and 3 days (Fig. 4A, B and C respectively) revealed a poor enrichment with non-specific cellular processes, indicating a similar uncontrolled gene expression as previously detected by RNA-seq data. On the other hand, the top enriched GO terms for the upregulated proteins of spheroids after the recovery period (day 10) revealed liver-related functions, such as lipid metabolic process, cholesterol transport, and plasma lipoprotein particle remodeling (Fig. 4D). Biological processes that were impaired by NaBut treatment include regulation of nuclear processes, such as RNA metabolism, transcription by RNA polymerase II, as well as chromosome and chromatin organization (Additional file 1: Fig. S4A and B). Although spheroids seem to reestablish its liver functions after the recovery period, some nuclear processes, including RNA metabolic process and chromatin organization, remain altered and might take longer to return to basal levels (Additional file 1: Fig. S4C). This can be explained by proteins having a slower turnover than histone acetylation. The full list of identified proteins is available as Additional file 5: Table S3.
Fig. 4
Liver function is impaired by chromatin decondensation and transcription activation. Gene ontology (GO) enrichment analysis of spheroids treated for A 1, B 2, C 3 days and D after recovery (day 10). Functional annotation was obtained using GOrilla [27]. Clustered network of the 200 upregulated proteins in spheroids treated with NaBut for 3 days E and F recovered spheroids (day 10). Clusters reflect connected portions of the network and correspond to functional categories of the proteins. Size of nodes represents p-value, color darkness represents the score (fold change enrichment times the p-value), and line thickness represents the score of interaction confidence retrieved from the software String (v11, https://string-db.org). The network was constructed by Cytoscape [20]
Proteins with the highest score in spheroids treated with NaBut for 3 days or spheroids recovered from treatment (day 10) were then used to construct a network of protein interactions (Fig. 4E and and F, respectively) to demonstrate the molecular and cellular mechanisms enriched in each of the conditions. Biological processes significantly enriched in treated spheroids revealed proteins mostly related to cellular response to stress and immune system, although proteins related to metabolism of lipids and proteins and signal transduction were also enriched (Fig. 4E). Interestingly, the pathways most enriched after spheroids are recovered from the treatment include post-translational protein phosphorylation, vesicle-mediated transport, and complement and coagulation cascades (Fig. 4F). Biological processes related to liver function, such as the TCA cycle, metabolism of lipids, and cholesterol metabolism, are also enriched once spheroids reestablish their homeostasis, indicating that the cells within 3D spheroids regulate their proteome toward recovering original liver functionalities. Interestingly, this regulation was not noticed in RNA-seq data, most likely due to the faster turnover of RNAs compared to proteins. In fact, by day 10 basically all RNA levels were back to baseline (Fig. 3A).
Altogether, our data show that NaBut treatment for 3 days induces histone hyperacetylation and, as an indirect effect, triggers transcriptional activation, most likely through chromatin decondensation. However, the proteome showed a slower regulation compared to histone modifications and mRNA, potentially contributing to re-establishing the baseline functionalities of spheroids prior to treatment. Enriched proteins related to lipid metabolic process, described above, are highlighted in the volcano plots for each of the treatment days (Fig. 5A). It is evident that most of these proteins are enriched on day 10, when the cells are no longer exposed to the treatment, indicating that cells returned their normal metabolism. On the other hand, proteins related to response to treatment (Fig. 5B) and transcription (Fig. 5C) are enriched specially on day 3, suggesting that the effect of NaBut treatment might be cumulative in the cells. Altogether, this indicates that our 3D cell culture is a suitable model for recovery studies as well as long-term treatments (model in Fig. 6A–B), approaches that are not supported by cells growing as a monolayer.
Fig. 5
NaBut treatment affects the proteome of spheroids. Volcano plots representing NaBut vs Nt fold change after treatment (days 1, 2, and 3) and recovery period (day 10). Highlighted bubbles display the relative abundance of proteins related to A lipid metabolic process, B cellular response to treatment, and C transcription
Fig. 6
Schematic representation of direct and indirect targets of NaBut treatment on spheroids. A NaBut treatment induces direct effects, such as histone hyperacetylation and indirect, such as chromatin decondensation and transcription activation. B Removal of treatment and reestablishment of standard culture condition revert the NaBut effects and cells return their normal physiology
留言 (0)