Cells and viruses
hACE2 expressing BHK-21 (BHK-21-hACE2) and Vero-E6 cells were provided by State Key Laboratory of Virology, Wuhan University. Vero-E6 cells were cultured in DMEM (Gibco, Cat.11995–040) supplemented with 10% fetal bovine serum (Gibco, Cat.10091–148) and 1% penicillin and streptomycin solution (BasalMedia, Cat.S110JV) at 37°C with 5% CO2. BHK-21-hACE2 cells were cultured in the above medium supplemented with 1 μg/mL puromycin (Beyotime, Cat.ST511) under the same conditions.
SARS-CoV-2 viruses (Wuhan strain, Beta, Delta, and Omicron variants) were amplified and titrated in Vero-E6 cells in DMEM (Gibco, Cat.11995–040) supplemented with 10% fetal bovine serum and 1% penicillin and streptomycin solution at 37°C with 5% CO2, and reported as TCID50/ml. The Delta variant (CSTR.16698.06. NPRC 6.CCPM-B-V-049-2105-8) used here is from National Pathogen Resource Center, NPRC and stocked at National Virus Resource Center. The South Africa variant (NPRC2.062100001) was obtained from National Institute for Viral Disease Control and Prevention, China CDC. All the viruses were propagated on the Vero-E6 cells.
Protein production and characterization
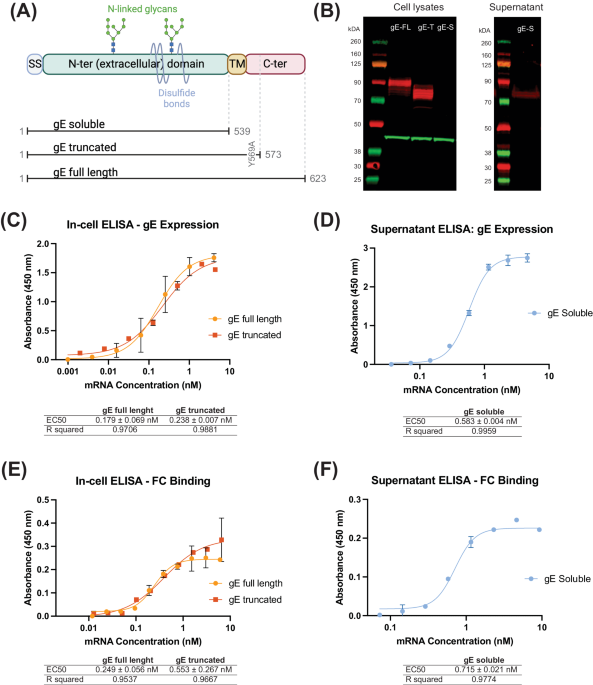
The variant S-trimer contains four hot spot mutations (K417N, E484K, N501Y and D614G), six stabilizing proline substitutions (F817P, A892P, A899P, A942P, K986P and V987P), a “GGSG” substitution at the furin cleavage site (residues 682-685), and a C-terminal T4 fibritin trimerization motif10. To produce the modified S-trimer, the codon optimized gene was synthesized and cloned into the mammalian expression vector SNT70. After transfection into Chinese hamster ovary (CHO) cells, the variant S-Trimer protein was purified from clarified supernatant as previously described. In brief, after the removal of CHO cells in the cell culture via depth-filtration (Cobetter), the clarified supernatant underwent half an hour of low pH treatment for viral inactivation (VI) at pH3.6. Then the resultant precipitates were removed and the pH was adjusted to neutral range. Three different chromatography steps were successively employed to remove host cell DNA, host cell proteins, and any other impurities, and finally nanofiltration was introduced as a preventative viral removal (VR) step.
SDS-PAGE and Size-Exclusion HPLC were run to check the purity and trimeric conformation of the variant spike protein. Biolayer interferometry (BLI) assays were performed on a GatorPrime (GatorBio) instrument at 30 °C with shaking at 1000 rpm, to determine the kinetic parameters and binding affinity of modified S-trimer to human ACE2. In general, the Fc-tagged human ACE2 (Sino Biological, Cat.10108-H02H) was immobilized to human Fc (HFC) Probes (Gator Bio, Cat.160003) at 10 μg/mL. The variant S-trimer was two-fold serially diluted in PBST prior to the measurement of the association and dissociation rates. The data were baseline subtracted prior to curve fitting performed using a 1:1 binding model. Kinetic parameters, kon and koff, which can be used to determine an equilibrium dissociation constant KD, were determined with a global fit applied to all data.
Animal studies
All animal studies were conducted following study protocols approved by the Institutional Animal Care and Use Committee (IACUC) of the National Center for Safety Evaluation of Drugs (NCSED).
Six to eight weeks old female BALB/c mice were purchased from the Laboratory Animal Management Department, Shanghai Institute of Family Planning Science and divided into different groups (ten mice each) and intramuscularly (i.m.) administered with a 1/10 human dose of experimental vaccine or control on day 0 and 21. The mice were bled on day 14 and day 35. Serum samples were collected and heat-inactivated at 56 °C for 30 min to evaluate antibody responses via ELISA and neutralization assays. On day 35, five mice in each group were sacrificed to assess T cell responses by intracellular cytokine staining (ICS) and ELISPOT.
The immunogenicity and challenge study in rhesus macaques was commissioned by the Wuhan Institute of Virology, Chinese Academy of Sciences. Twelve rhesus macaques (weighing 5–7 kg) of 5–7 years old were randomly divided into 2 groups: variant vaccine or adjuvant control. Groups of 6 macaques were (three males and three females) intramuscularly immunized with a full human dose of the vaccine or adjuvant at day 0 and day 21. Blood was collected and sera were prepared on day -1, day 8, day15, day 21, day 29, day 35 and day 43 to detect antibody responses. On day 35, the PBMCs were separated to evaluate vaccine induced cellular immune responses by ICS and ELISPOT. On day 65, all rhesus macaques were transferred to Biosafety Level 4 Laboratory (BSL-4) in Wuhan National Biosafety Laboratory, Chinese Academy of Sciences and challenged with 1 × 105 TCID50 SARS-CoV-2 (2019-nCoV-WIV04, GISAID: EPI_ISI_402124) by tracheal intubation. After the viral challenge, body weight and temperature were monitored daily, and the swabs of throat, nasal, anal and blood were daily collected to detect changes of viral load. On day 72, rhesus macaques were euthanized for detecting viral loads of lungs and trachea-bronchus by qRT-PCR and evaluating lung histopathology by hematoxylin and eosin (H&E) staining.
The immunogenicity and challenge study in Golden Syrian hamsters was commissioned by the Institute of Medical Laboratory Animals, Chinese Academy of Medical Sciences. Hamsters were intramuscularly immunized twice on Day 0 and Day 21. On Day 35, blood from all hamsters was collected to detect antibody responses. On Day 35, all hamsters were challenged intranasally with 105 TCID50 SARS-CoV-2 (Beta or Delta). After viral challenge, the body weights and temperature of the hamsters were monitored daily. On Day 49, hamsters in each group were euthanized for detecting viral loads of lungs and nasal turbinates by qRT-PCR and for evaluating lung histopathology by H&E staining.
Enzyme-linked immunosorbent assay (ELISA)
Binding antibodies were assessed by ELISA. Briefly, 96-well plates (Corning, Cat.9018) were coated at 4 °C overnight with 1.0 mg/mL S protein in phosphate-buffered saline (PBS). Plates were washed once with PBST (0.05% Tween 20 in PBS) and blocked with 5% milk-PBS for 2 h at 37 °C. Five-fold serial dilutions of heat-inactivated serum were made in 2% milk-PBS for 100 μL/well and plates were incubated at 37 °C for 1 h. This was followed by 1 h incubation at 37 °C with a dilution of horseradish peroxidase (HRP) conjugated Goat anti-mouse IgG (1:10000) (Bio-Rad, Cat.1706516), Goat anti-hamster IgG (1:15000) (Invitrogen, Cat.PA1-28823), or Goat anti-monkey IgG (1:5000) (BETHYL, Cat.A140-102P). Plates were then washed six times, 100 μL per well of TMB substrate system solution (SeraCare, Cat.5120–0038) was added for 15 min in the dark and stopped by adding 50 μL 2 M sulfuric acid. The absorbance was measured as OD450-OD620 using a microplate reader (Molecular Devices, SpectraMax iD3).
The endpoint titers were defined as the highest reciprocal dilution of serum to give an absorbance >2.1-fold of the background values that defined as 0.05. Antibody titer below the limit of detection was determined as half the limit of detection.
Pseudovirus-based neutralization assay
The SARS-CoV-2 pseudoviruses were prepared using a replication-deficient VSV-based rhabdoviral pseudotyping system expressing firefly 485 luciferase (VSV-dG-fluc), which was obtained from State Key Laboratory of Virology, Wuhan University. Pseudovirus production has been described in another study of our team10. For neutralization assay, 60 μL 4-fold serially diluted inactivated serum and 60 μL 800 TCID50 SARS-CoV-2 pseudoviruses were added into 96-well plates (Corning, Cat.3599) one after another. The mixture was incubated at 37 °C for 1 h, and then added to BHK-21-hACE2 cells in a 96-well white plate with clear bottom (Corning, Cat.3610). Luciferase activity was measured 24 h later using luciferin-containing substrate (PerkinElmer, Cat.6066769). The nAb titer was calculated by the dilution number of 50% inhibition condition. The neutralizing titer was calculated according to Reed-Muench Method.10
Live virus-based neutralization assay
All SARS-CoV-2 live virus-related experiments were conducted in the BSL-3 laboratory, the macaque serum samples were measured by plaque reduction neutralization test (PRNT) in the State Key Laboratory of Virology, Wuhan University and the hamster serum samples were measured by cytopathic effect (CPE) in the Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences. The CPE neutralization assays were performed as previously described of our team10. Briefly, medium containing serum at varying dilutions in 96-well plates was pre-incubated with an equal volume of live SARS-CoV-2 solution diluted to contain 100 TCID50. After neutralization in a 37 °C for 1 h, the mixture was added to Vero E6 cells in a 96-well plate. After 3–5 days’ incubation at 37 °C, cytopathic effect (CPE) of each well was recorded under microscopes, and the neutralizing titer was calculated by the dilution number of 50% protective condition.
For PRNT50 assay, sera were three-fold serially diluted and incubated with an equal volume of 1000 plaque-forming units (pfu) /mL virus at 37 °C for 1 h. The serum–virus mixture was added to Vero E6 cell monolayers seeded in 24-well plates and incubated at 37 °C for 1 h. The inoculum was removed and replaced with 1.0 mL 0.9% methylcellulose complete DMEM. Plates were incubated at 37 °C for 96 h and fixed with 8% paraformaldehyde for 30 min at room temperature. Following fixation, plates were replaced the fixed solution with 0.05% crystal violet for staining. The number of plaques per well was counted and the virus neutralizing antibody titer FRNT50 defined as the highest serum dilution resulting in 50% reduction of plaques.
Intracellular cytokine staining (ICS) assay
The splenocytes or PBMCs were harvested from immunized animals on day 14 after the second immunization and analyzed by flow cytometry. Splenocytes stimulated with Phorbol 12-myristate13-acetate (Sigma-Aldrich, Cat.79346) and Ionomycin (Yeasen, Cat.50401ES03) served as positive controls, unstimulated cells were used as negative controls. S spike peptide pool from original strain (Wuhan-Hu-1), containing 170 peptide counts (synthesized by GenScript, China)10,45, each containing 18 amino acids in length with 7 amino acids offset and 11 amino acids overlapped (Supplementary table. 1). A total of 1 × 106 splenocytes were stimulated with 1 μg/mL S peptide pool in the presence of 1 μg/mL Brefeldin A (BD, Cat. 555029) and 1 μg/mL Anti-CD28 (Mabtech, Cat.FS-2122-10) at 37 °C for 6 h. Cells were stained with LIVE/DEAD Aqua and a panel of flow cytometry antibodies specific for cell surface markers: PB anti-mouse CD3 (BioLegend, Cat.100214), FITC anti-mouse CD4 (BD, Cat.553047) for mouse study or PB anti-human CD3 (BD, Cat.558124), FITC anti-human CD4 (BD, Cat.550628) for NHP study at 2–8 °C for 30 min. Following washing and permeabilization (BD, Cat.554714), cells were further stained with flow cytometry antibody mixture: APC anti-mouse IFN-γ (BD,Cat.554413), PE-Cy7 anti-mouse IL-2 (BD, Cat.560538), PE anti-mouse TNF-α (BD, Cat.554419) for mouse study or APC anti-human IFN-γ (BD, Cat.554702), PE-Cy7 anti- human IL-2 (BD, Cat.560707), PE anti- human TNF-α (BD, Cat.557068) for NHP study at 2–8 °C or 30 min. The stained cells were analyzed using a BD FACS Canto II flow cytometer using BD FACS Diva software.
Enzyme-linked immunospot (ELISPOT) assay
The splenocytes or PBMCs were separated from immunized animals on day 14 after the second immunization and resuspended in complete RPMI-1640 supplemented with 10% FBS. For in vitro stimulation, a total of 1 × 106 splenocytes were pre-incubated with 0.2 ug/mL CD28 to stimulated and incubated with S peptide pool in complete RPMI-1640 for 72 h at 37 °C. Splenocytes stimulated with ConA (Sigma-Aldrich, Cat.L7647-25MG) served as positive controls. The cytokines were detected by Monkey IFN-γ/IL-2/TNF-α FluoroSpotPlus Kit (Mabtech, Cat.FSP-212822-10), Human IL-4/IL-5 FluoroSpotFLEX Kit (Mabtech, Cat.X-16B08W-1) or Mouse IFN-γ/IL-2/TNF-α FluoroSpot FLEX Kit (Mabtech, Cat.X-41A42B45W-10), respectively, according to the manufacturer’s instructions. Plates were scanned and counted on a ELISPOT reader (CTL, S6).
Viral RNA assay
SARS-CoV-2 RNA levels were monitored by qRT-PCR. Briefly, RNA, as template of qRT-PCR, was extracted from swabs and organ tissue homogenates by QIAamp Viral RNA Mini Kit (Qiagen) according to the manufacturer’s instructions. RNA copies were measured by HiScript® II One Step qRT-PCR SYBR® Green Kit (Vazyme Biotech Co., Ltd). The forward and reverse primers targeting SARS-CoV-2 spike protein (S) gene for RBD-qF1 was 5'-CAATGGTTAAGGCAGG-3' and for RBD-qR1 was 5'-CTCAAGGTCTGGATCACG-3'. qRT-PCR was conducted by an ABI StepOne under the following reaction conditions: (1) 50 °C for 3 min; (2) 95 °C for 30 s; (3) 40 cycles of 95 °Cfor 30 s; (4) 60 °C for 30 s.
H&E staining and immunohistochemistry
To evaluate lung histopathology by H&E staining, lung tissues from rhesus macaques and hamsters were fixed by 10% buffered formalin and processed for paraffin embedding. Paraffin blocks were cut into 5 μm sections and stained with hematoxylin and eosin. Lung pathology, including overall lesion extent, pneumocyte hyperplasia, and inflammatory infiltrates, was assessed and classified into three types: mild, moderate and severe lung inflammation.
Statistical analysis
All statistical analyses were performed using GraphPad Prism 8.0. Antibody titers and viral loads were log10 transformed prior to statistical analysis. All blots derive from the same experiment and were processed in parallel. Statistical significance was determined using ANOVA for multiple comparisons. Student’s t-tests were applied to compare two groups. And correlations were assessed by two-sided Spearman rank correlation tests, three important parameters from the correlates analysis shown as R-value, P-value and 95% confidence intervals, and shaded areas represent 95% confidence intervals. Values of p < 0.05 are considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.

留言 (0)