
記住我

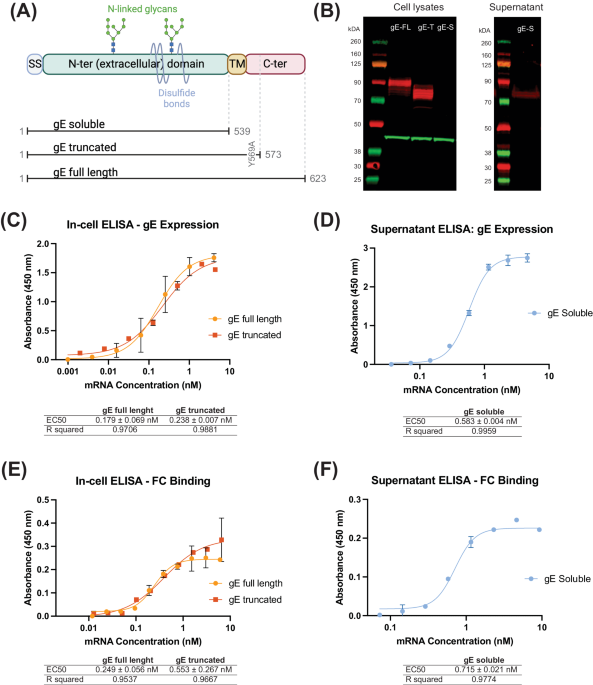
Since glycoprotein E (gE) is the most abundantly expressed antigen both on the surface of the varicella zoster virus (VZV) particles and on the infected cells24 and is used in the currently approved subunit vaccine Shingrix®16, we therefore designed our mRNA constructs to encode this protein. gE is a transmembrane protein consisting of a long N-terminal extracellular domain expressed on the cell or viral surface, a short transmembrane domain anchoring the protein to the viral or host cell membrane, and a short C-terminal cytoplasmic domain. The N-terminal extracellular domain mediates its binding to insulin-degrading enzyme (IDE) on the cell surface26 and represents the main VZV antigen for recognition by antibodies and T-cells. The C-terminal domain has been shown to play a critical role in gE protein localization, however, the relationship between localization of the membrane protein and immunity is poorly understood. Therefore, our experimental designs for selection of a particular gE antigen-encoding mRNA candidate were based on comparison of the full-length gE antigen to its truncated variants. Specifically, we designed three versions of the gE antigen (Fig. 1A): (i) full-length gE (gE full-length), as present on the surface of the virion or naturally infected cells24, (ii) truncated gE (gE truncated), generated by deleting a part of the C-terminal domain and including a single amino acid substitution (Y569A) altering one of the trans-Golgi network (TGN) localization motifs, as described previously25, and (iii) soluble gE (gE-soluble), which is expressed as the extracellular domain of the protein and similar to the antigen sequence in Shingrix®. In truncated gE, the C-terminal domain containing the trans-Golgi network (TGN) localization motif was modified by a single point-mutation: AYRV to AARV (Y569A). These modifications were made in truncated gE with the aim to address gE trafficking and thus potentially improve the surface expression of gE after translation. In addition to a codon-optimized open reading frame, the mRNA also contained a proprietary transcription initiation sequence (TIS), human β-globin (HBG) 5′ UTR and 3′ UTR sequences, and a polyA tail sequence. The final transcript was 5′ capped and contained N1-methylpseudouridine (m1Ψ) modified bases.
Fig. 1: Design and in vitro characterization of mRNA constructs expressing the three different versions of Varicella Zoster Virus (VZV) glycoprotein E (gE).
A Schematic representation of the various domains of VZV gE. The N-terminal domain contains disulfide bonds, N-glycans, and O-glycans (O-glycans not shown). Three versions of the VZV gE protein that we designed are shown: The soluble gE protein (gE soluble) contains only the signal sequence (SS) and the extracellular N-terminal domain. The truncated gE protein (gE truncated) contains the signal sequence, extracellular N-terminal domain, transmembrane (TM) domain and part of the C-terminal domain with one point mutation in the trans-Golgi network (TGN) localization motif, wherein AYRV sequence was modified to AARV (Y569A). The full-length gE protein (gE full-length) contains the entire wild-type open reading frame of the glycoprotein. B Western blot of cell lysates (left) and cell supernatants (right) after transfection with the indicated gE-expressing mRNA constructs. gE-FL – gE full-length, gE-T – gE truncated, and gE-S–gE soluble protein. Red bands correspond to the gE protein, green bands correspond to beta actin that is used as loading control. The observed molecular weight sizes for gE-FL (90 kDa) and gE-T (80 kDa) correspond to their partially glycosylated forms. gE-S (observed at 75 kDa) is not detected in the cell lysates, as expected, but is instead detected in the supernatant (right). C, D Detection of gE expression on cell surface (gE full-length and gE truncated) and in supernatant (gE soluble) by ELISA. EC50 values (in nM) are shown as Mean ± SD; R square values are shown to highlight the goodness of fit. The detection is performed using anti-gE antibodies. E, F Binding of human Fc fragment to cell surfaces expressed gE (gE full-length and gE truncated) (E) and gE in supernatant (F) to demonstrate the appropriate conformation and functional nature of the gE proteins. EC50 values (in nM) are shown as Mean ± SD; R square value is shown to highlight the goodness of fit.
After generation of the mRNA constructs, we successfully demonstrated the expression of all three gE variants in transfected HEK293FT cells by Western blotting (Fig. 1B) and in transfected HeLa cells by in-cell ELISA (for gE full-length and gE truncated; Fig. 1C) and supernatant ELISA (for gE soluble; Fig. 1D). The various gE proteins migrated on the PAGE gel at expected molecular weights with a clear difference between the sizes of the full length and truncated (gE truncated and gE soluble) versions of the protein. As expected, the soluble gE was not detected in cell lysates but was detected in the supernatants of transfected cells both by Western blot (Fig. 1B) and by ELISA (Fig. 1D). In-cell ELISA (gE full-length and gE truncated) and ELISA (gE soluble) showed that the expression levels of full-length gE and truncated gE antigens (Fig. 1C) were similar to the soluble gE antigen (Fig. 1D). This was further confirmed when HEK293FT cells, expressing the three gE antigen variants, were treated with Brefeldin A and we observed similar expression levels via Western blot (data not shown). Additionally, we characterized the functionality of the expressed gE proteins by their ability to bind human Fc. To that end, we developed Fc-binding ELISAs and observed that all three gE variants bound the recombinant human Fc similarly (Fig. 1E for gE full-length and gE truncated antigens binding to Fc; Fig. 1F for gE soluble antigen binding to Fc). These results confirmed the successful designs of the mRNA constructs encoding the three gE antigen variants, their appropriate expression upon transfection, and demonstration of functional antigenic conformation via human Fc-binding.
Production of mRNAs and mRNA-LNP vaccine candidates for in vivo studiesTo generate vaccine candidate materials for the in vivo studies, the three gE-encoding (gE full length, gE truncated and gE soluble protein) mRNAs were produced by in vitro transcription, purified using lithium chloride precipitation, and stored at ≤−65 °C until further use. The purified mRNAs were tested for the following key attributes: concentration, identity, purity/integrity, poly(A) tail length, % capping efficiency, % N1-methyl-pseudouridine (m1Ψ) incorporation, and levels of various impurities (e.g., RNase, endotoxin, E.coli DNA and residual NTPs). These analytical test results are shown in Supplementary Table S2. Thereafter, all three purified gE-encoding mRNAs were encapsulated in either NOF-lipid-based LNPs or SM102-lipid-based LNPs and the final formulated mRNA-LNPs were tested for the following critical attributes: Particle size, Polydispersity index (PDI), % mRNA encapsulation, % mRNA purity post encapsulation and mRNA concentration. The analytical test results of the six mRNA-LNPs are shown in Supplementary Table S3.
Assessment of in vivo delivery and preliminary immunogenicity of GLB mRNA-LNP vaccine candidatesWe aimed to evaluate immunogenicity of the three gE-encoding GLB mRNA-LNP vaccine candidates in mice. However, before evaluating all three gE variants, we wanted to conduct a preliminary study using the mRNA-LNP formulation encoding the full-length glycoprotein E (gE). The LNP formulation was based on proprietary cationic lipids from NOF corporation. The aim of this preliminary study was to quickly test if the novel NOF-LNP formulation was effective in delivering the mRNA in vivo, using the gE full-length antigen, before conducting the full comparative evaluation of the gE variants. As positive control, the currently approved subunit vaccine Shingrix® was used.
To mimic pre-existing immunity against VZV, which is expected to be present in dormant state in the target human population, C57BL/6 female mice were primed with a full human dose (500 μL) of the live-attenuated Varicella Zoster virus (Varivax®) by subcutaneous administration. It should be noted that the live-attenuated VZV (Varivax®) cannot replicate in mouse cells and establish latency in this animal. Mice were immunized with two intramuscular injections of the gE full-length encoding mRNA-LNP (NOF) candidate, 4- and 8-weeks post Varivax® priming, as shown in Fig. 2A. Immunizations were performed at a dose of 5 μg per animal, in a total volume of 50 µL per mouse (25 µL in each hind limb). Mice injected with Varivax® only or saline were included as controls.
Fig. 2: Initial immunogenicity of mRNA-LNP vaccine encoding for the VZV full-length gE (gE full-length) in mice.
Female C57BL/6 mice were primed with a full human dose of the live attenuated VZV vaccine (Varivax®) by subcutaneous administration. Mice were then injected intramuscularly with 5 μg of mRNA-LNP vaccine candidates, expressing VZV gE full-length, or saline (Varivax® only group) at weeks 4- and 8- post Varivax® administration. NOF-LNP formulated mRNA, delivering gE full-length, is the test candidate while Shingrix® is used as a positive control. A Scheme of immunizations and sample collections schedule. B End-point titers of gE-specific IgG binding antibodies detected in mice at the indicated timepoints and evaluated by ELISA. Arrows indicate the two mRNA-LNP/Shingrix® immunization timepoints. Data shown as Mean ± SEM, and is representative of two independent experiments using 10 animals per group. Y-axis represents Log10 end-point titers; X-axis represents weeks post-Varivax® prime. C Antigen-specific effector T cell responses measured from whole blood using a murine IFN-γ/IL-2 Double-Color Enzymatic ELISpot Assay. Data shown as Mean ± SEM. Y-axis represents Spot Forming Cells (SFCs) per million peripheral blood cells. Each SFC corresponds to an effector T cell secreting either one or both cytokines in response to stimulation by the gE overlapping peptides pool.
Anti-gE IgG binding antibodies were detected by ELISA from serum samples collected at 0-, 4-, 6-, 8-, and 10-weeks post Varivax® prime. Immunization of mice with 5 μg of gE full-length mRNA-LNP (NOF) induced high levels of anti-gE serum IgG binding antibodies that were comparable to those elicited by Shingrix®, as shown in Fig. 2B. Anti-gE binding antibody titers observed in animals immunized with mRNA-LNP (NOF) vaccine increased significantly after each immunization, when compared to animals primed with Varivax® only.
Effector T cell responses were assessed from peripheral blood 2 weeks post Varivax® prime (baseline response), and 1 week post the first and second mRNA-LNP immunizations using the murine IFN-γ/IL-2 Double-Color Enzymatic ELISpot Assay. gE-specific activated T cells secreting either IFN-γ or IL-2 or both cytokines, after re-stimulation with gE overlapping peptides, were quantified (Fig. 2C). At baseline, the number of spot-forming cells (SFC) in all groups were similarly low, below 10 SFC per million blood cells. However, following two immunizations with 5 μg of mRNA-LNP (NOF) vaccine candidate, all animals developed strong gE-specific effector T cell responses that were higher than those observed in mice administered with two injections of Shingrix®. No long-lasting T-cell response was detected in animals primed with Varivax® alone. Taken together, the data from this preliminary study shows that the GLB mRNA-LNP (NOF) formulation is successful in delivering the gE glycoprotein and successful in eliciting a strong immunogenic response, comparable to Shingrix®, in mice.
Assessment of comparative immunogenicity of GLB mRNA-LNP vaccine candidates encoding the three different versions of gE antigenBased on the success of the above preliminary in vivo study in mice, we designed this expanded comparative immunogenicity study to evaluate all three mRNA-LNP vaccines candidates, encoding the three different versions of the gE antigen, and to compare them to Shingrix®. For this study, the three gE antigen variants-encoding mRNAs were formulated in two different LNP formulations for in vivo testing: one using the proprietary NOF LNP and the other using SM102 LNP, as a LNP comparator. SM102 LNP was chosen as a comparator because successful delivery of mRNA encoding for VZV’s gE by SM102 LNP has been reported earlier by Monslow et al.27. Each mRNA-LNP candidate was produced, tested for critical attributes (mRNA content, mRNA integrity and purity, LNP size and polydispersity) and thereafter frozen at <−65 °C until use in the study. In this study, two different antigen doses for each of the gE antigens were tested: a high dose of 5 µg and a low dose of 1 µg.
Consistent with the previous mouse study (Fig. 2A), this study in C57BL/6 female mice was also similarly designed. All mice received a full human dose (500 μL) of the live attenuated Varicella vaccine (Varivax®) by subcutaneous administration to set the initial VZV infection. 4 weeks post Varivax® priming, mice were administered with two intramuscular injections of the formulated mRNA, 4 weeks apart (Fig. 3A). Additionally, one group of mice was administered with 5 µg of Shingrix® (1/10 of the human dose) twice, 4 weeks apart, as positive control or an active comparator. Vaccines were administered in a total volume of 50 µL per mouse (25 µL in each hind limb). Mice injected with Varivax® only and saline were included as (negative) controls.
Fig. 3: Comparative immunogenicity of mRNA-LNP vaccine candidates encoding three different versions of VZV gE antigen.
Female C57BL/6 mice (5 animals per group) were primed with a full human dose of the live-attenuated VZV) vaccine (Varivax®) by subcutaneous administration. Mice were then injected intramuscularly with 1 or 5 μg of mRNA-LNP vaccine candidates, encoding the three gE variants, or saline (Varivax® only group) at 4 and 8 weeks post Varivax® administration. A Scheme of immunization and sample collection schedule. B Longitudinal Log10 end-point titers (Y-axis) of gE-specific IgG binding antibodies detected in sera collected from mice at the indicated timepoints and evaluated by ELISA. Arrows indicate mRNA-LNP/Shingrix® immunization time points. Data shown as Mean ± SEM. C Quantitative assessment of anti-gE serum IgG binding antibodies in mice from all vaccinated groups at 12 weeks post Varivax® prime, assessed by ELISA and expressed as area under the curve (AUC) (Y-axis) based on absorbance values. Data shown as Mean ± SEM.
Anti-gE binding antibodies were detected by ELISA from sera of mice collected on 0, 4, 8, 12, 16, 20, and 24 weeks. Serum IgG binding antibody levels induced by NOF LNPs encapsulating 5 µg of mRNA were equivalent to antibody levels observed in animals vaccinated with Shingrix® (Fig. 3B). Moreover, NOF LNP-formulated vaccine induced higher levels of gE-specific binding antibodies when compared to SM102 LNP-formulated vaccines in a dose-dependent manner (Fig. 3C). The peak binding antibody response was observed at week 12 i.e., 4 weeks post 2nd vaccination (Fig. 3B, C). As expected, mice vaccinated with saline had no detectable anti-gE serum IgG throughout the study.
Previous studies suggest that CD4+ T cells play a critical role in protecting against reactivation of VZV28,29. The decline in VZV-specific CD4+ T-cell responses has been associated with an increased risk of herpes zoster in older adults30,31. Moreover, immunocompromised individuals with impaired CD4+ T-cell function, such as people living with HIV, but not antibody deficiencies, are at a higher risk of developing herpes zoster and are more likely to experience severe and prolonged episodes28. Therefore, vaccines that can boost CD4+ T cell responses are believed to provide additional protection against reactivation of VZV. To that end, to assess amplitude and kinetics of T cell responses induced by the various mRNA-LNP vaccines encoding the three gE antigen variants, peripheral blood was collected 2 weeks post Varivax® prime, and at 1 and 2 weeks post each vaccination (mRNA-LNP or Shingrix®), as depicted in Fig. 3A. Peripheral blood mononuclear cells were stimulated with a gE overlapping peptide pool, and the number of gE-specific cells secreting IFN-γ and/or IL-2 was quantified by dual-color ELISpot. Priming with Varivax® induced very weak T-cell responses in mice (Fig. 4A), as also previously described by others19,27. Effector T cell responses peaked at day 7 post each vaccination, either by the mRNA-LNP vaccines or the active control (Shingrix®). Overall, the mRNA-LNP vaccine expressing the soluble version of gE performed poorly when compared to mRNA-LNP vaccines encoding the full-length gE or the truncated gE; regardless of the type of LNP (NOF vs. SM102) formulation used (Fig. 4A, B). Remarkably, the three NOF LNP-formulated vaccines encoding each of the three gE antigen variants, induced very high levels of poly-functional effector T cells, evidenced by secretion of more than one cytokine simultaneously, which were higher than levels induced by the Shingrix® vaccinated animals.
Fig. 4: mRNA-LNP vaccine candidates, encoding VZV gE, induces strong antigen-specific T cell responses.
A, B Female C57BL/6 mice (5 animals per group) were primed with a full human dose of the live attenuated VZV vaccine (Varivax®) by subcutaneous administration. Mice were injected intramuscularly 5 μg of the mRNA-LNP vaccine candidates, expressing the three gE variants, or Shingrix® at 4 and 8 weeks post Varivax® administration. A, B Antigen-specific effector T cell responses were measured from whole blood cells collected at week 5 i.e., 1 week post 1st immunization (A) or week 9, i.e., 1 week post 2nd immunization (Busing a murine IFN-γ/IL-2 Double-Color Enzymatic ELISpot Assay. Data shown as Mean ± SEM. Y-axis in both panels, A and B, represents Spot Forming Cells (SFCs) per million peripheral blood cells. Each SFC corresponds to an effector T cell secreting either one or both cytokines in response to the gE overlapping peptides pool.
To bolster the above findings, we conducted an additional study to measure effector T cell responses from splenocytes, collected 1 week post 2nd immunization with mRNA-LNP (NOF), mRNA-LNP (SM102) or Shingrix®, by intracellular cytokine staining (ICS). In this study, mice were administered with two intramuscular injections of the LNP (NOF or SM102) formulated mRNA or Shingrix®, 4 weeks apart, without Varivax® priming (Fig. 5A). Mice injected with saline were included as (negative) controls. Splenocytes obtained a week after the second immunization were stimulated with gE overlapping peptide pool, and the number of gE-specific CD4+ or CD8+ T cells secreting IFN-γ, TNF-α and/or IL-2 were quantified by flow cytometry (Supplementary Figure S1). The gE-specific effector T cell responses were predominantly CD4+ T cell-mediated, since less than 0.5% of the CD8+ T cells in the spleen detected were secreting cytokines upon stimulation with gE peptides (Fig. 5B and Supplementary Figure S2A). Administration of NOF or SM102 formulated mRNA-LNP vaccine candidates induced potent CD4+ T cell responses, as seen by the number of gE-specific CD4+ T cells secreting either one, two or all three of the cytokines IFN-γ, TNF-α and IL-2 (Fig. 5C and Supplementary Figure S2B). Importantly, both NOF and SM102-formulated mRNA-LNP vaccines induced higher levels of CD4+ T cell responses than Shingrix®.
Fig. 5: CD4+ T cell-biased cellular response induced by gE mRNA-LNP vaccines.
A Animals were divided in groups of 7, and subsequently immunized with 2 intramuscular doses, 4 weeks apart, of gE mRNA-LNP (NOF or SM102) or Shingrix®. Mice injected with saline were included as negative controls. One week after the last immunization, spleens were harvested and stimulated with overlapping peptide pools from VZV gE protein, and percentage of CD8+ (B) and CD4+ (C) T cells producing IFN-γ, TNF-α, and IL-2, was measured by flow cytometry.
Evaluation of immunological memory induced by GLB mRNA-LNP vaccine candidatesEffective vaccines are expected to generate adaptive immunological memory in the B cell (BMEM) and T cell (TMEM) memory compartments. This memory is crucial for rapid recall of the immune responses to avoid re-activation of VZV and causation of herpes Zoster disease27,32. To assess if vaccination with the gE antigen encoding mRNA-LNP vaccine candidates could lead to long-term persistence of humoral responses, the presence of gE-specific long-lived plasma cells was investigated in the bone marrows and effector memory CD4+ T cells in the spleens of mice, collected 16 weeks post 2nd immunization, as depicted in schematics on Fig. 3A. gE-specific long-lived plasma cells (LLPCs), quantified as the number of bone marrow-derived anti-gE antibody (IgG) secreting cells (ASC) by B cell ELISpot assay, induced by 5 µg of mRNA formulated with NOF LNP, were comparable to responses observed in animals vaccinated with Shingrix® (Fig. 6A). Additionally, gE-specific CD4+ effector memory T cells, characterized as CD4+ CD44+ CD62L-, were quantified from spleens. We observed that gE-specific effector memory CD4+ T cell responses, induced by 5 µg of mRNA formulated with NOF LNP, were comparable to responses observed in animals vaccinated with Shingrix® (Fig. 6B). Cumulatively, based on the overall gE-specific antibody titers, antigen-specific T cell responses and LLPCs and T cell memory results, we selected the gE full-length for future studies to support clinical development of this vaccine candidate.
Fig. 6: mRNA-LNP vaccine candidates, encoding VZV gE, induces antigen-specific LLPCs and T (TMEM) cell memory responses in mice.
C57Bl/6 mice (5 animals per group) were primed with live attenuated VZV vaccine (Varivax®), and vaccinated twice, 4 weeks apart, with the mRNA-LNP vaccine candidates or Shingrix®. Immunological memory was evaluated 16 weeks post last immunization. A Long lived plasma cells in the bone marrow secreting antibodies that bind to VZV gE, quantified by B cell ELISpot assay. Y-axis represents Spot Forming Cells (SFCs) per million bone marrow-derived cells. B Percentages of effector memory CD4+CD44+CD62L- T cells (Y-axis) secreting either IFN-γ, TNF-α, and IL-2 alone or any of the two or all three cytokines. Data shown as Mean ± SEM.
Assessment of pro-inflammatory cytokines induced by GLB mRNA-LNP Shingles vaccine candidatesDespite the high efficacy of Shingrix®, vaccinees have reported severe local and systemic reactions post vaccination21,22. Thus, aware of the pronounced reactogenicity of Shingrix®, one of our major goals was to develop a mRNA-LNP vaccine against herpes zoster that is at least comparable with respect to immunogenicity of Shingrix® but with potentially lower induction of systemic inflammation.
To assess systemic inflammation induced by the selected gE full-length mRNA-LNP vaccine candidates, pro-inflammatory cytokines were measured from sera collected before immunization and 6 h and 24 h after the 1st immunization with mRNA-LNP (NOF) formulated vaccine candidate or mRNA-LNP (SM102) formulated vaccine candidate or Shingrix®, using the LEGENDplex™ Mouse Anti-Virus Response Panel. Interestingly, both NOF and SM102 lipid-formulated mRNA-LNP vaccines induced significatively lower levels of pro-inflammatory cytokines such as MCP-1 (Fig. 7A), CXCL-1 (Fig. 7B), and CXCL-10 (Fig. 7C) post vaccination than Shingrix®. Other pro-inflammatory cytokines such as interferons, IL-1β and IL-6 (that were included in the LEGENDplex™ Mouse Anti-Virus Response Panel) were not detected at any of the time points across all groups. These findings suggest that additional investigations will be necessary in future to allow for the full assessment of local and systemic toxicity of our mRNA-LNP in comparison to Shingrix®.
Fig. 7: mRNA-LNP vaccine candidates, encoding VZV gE, induce lower levels of inflammatory cytokines in mice than Shingrix®.
Female C57BL/6 mice (7 animals per group) were immunized with 5 µg of either SM102 LNP-formulated mRNA encoding gE full-length or NOF LNP-formulated mRNA encoding gE full-length or Shingrix® or saline alone (negative control). Pro-inflammatory cytokines (MCP-1 in panel A, CXCL1/KC in panel B, and CXCL10/IP-10 in panel C) were quantified in serum of immunized animals pre-dose, 6 h and 24 h post dosing, using LEGENDplex™ Mouse Anti-Virus Response flow-based multiplexed assay. Y-axis denotes the cytokine concentrations in pg/mL. Data shown as Mean ± SEM. Two-way Analysis of Variance (ANOVA) with Tukey’s multiple comparison test was performed to determine statistical significance. *p < 0.05, **p < 0.005, ***p < 0.0005, ****p < 0.0001; ns - not significant. MCP-1 monocyte chemoattractant protein-1, CXCL1/KC chemokine (C-X-C motif) ligand 1/keratinocyte-derived chemokine, CXCL10/IP-10 chemokine (C-X-C motif) ligand 10/ IFN-γ-inducible protein 10.
留言 (0)