
記住我
Microglia are the resident immune cells of the brain (Kreutzberg, 1996; Tremblay et al., 2011). They are derived from primitive myeloid progenitors which colonize the neuroepithelium during the early embryonic development (Ginhoux and Prinz, 2015). Since their discovery by Río-Hortega in 1919, the progress in bioengineering and pharmacology have been pivotal to investigate microglia functions. The 21st century has been the golden age of microglia research thanks to the development of selective ligands and antibodies along with transgenic mouse lines that allowed the visualization and manipulation of microglia (Sierra et al., 2019). These approaches revealed a critical role of microglia in response to infection and injury (Herz et al., 2017) and an active role in brain development and adult physiology (Tremblay et al., 2011). Many research reports are indeed focused on the role of microglia in shaping synaptic plasticity, sculpting brain circuits and in doing so modulating learning and memory processes and behavior (Tremblay and Majewska, 2011; Miyamoto et al., 2013; Morris et al., 2013; Tay et al., 2017; Taylor et al., 2017; Andoh and Koyama, 2021; Cornell et al., 2022).
Overall, the goal of this review is to gain new insights into the causal role of microglia in modulating neural plasticity, learning and memory and behavior via loss-of-function strategies.
We first described the methods that have been used to deplete microglia and their limitations (section 2), which are critical to interpret the available data regarding microglia function. Second, we described the effect of depleting microglia on structural and functional synaptic plasticity in the adult brain (section 3). Third, we summarized the effects of microglia depletion on learning and memory and behavior (section 4).
Our review does not cover the signaling pathways involved in the interaction between microglia and neurons, nor the molecular mediators of microglia responses and the various effectors released by microglia to modulate synaptic function or the role of microglia in regulating plasticity during brain development. We refer readers to excellent reviews on these topics (Miyamoto et al., 2013; Schafer et al., 2013; Frost and Schafer, 2016; Neniskyte and Gross, 2017; Werneburg et al., 2017; Marinelli et al., 2019; Cserép et al., 2021).
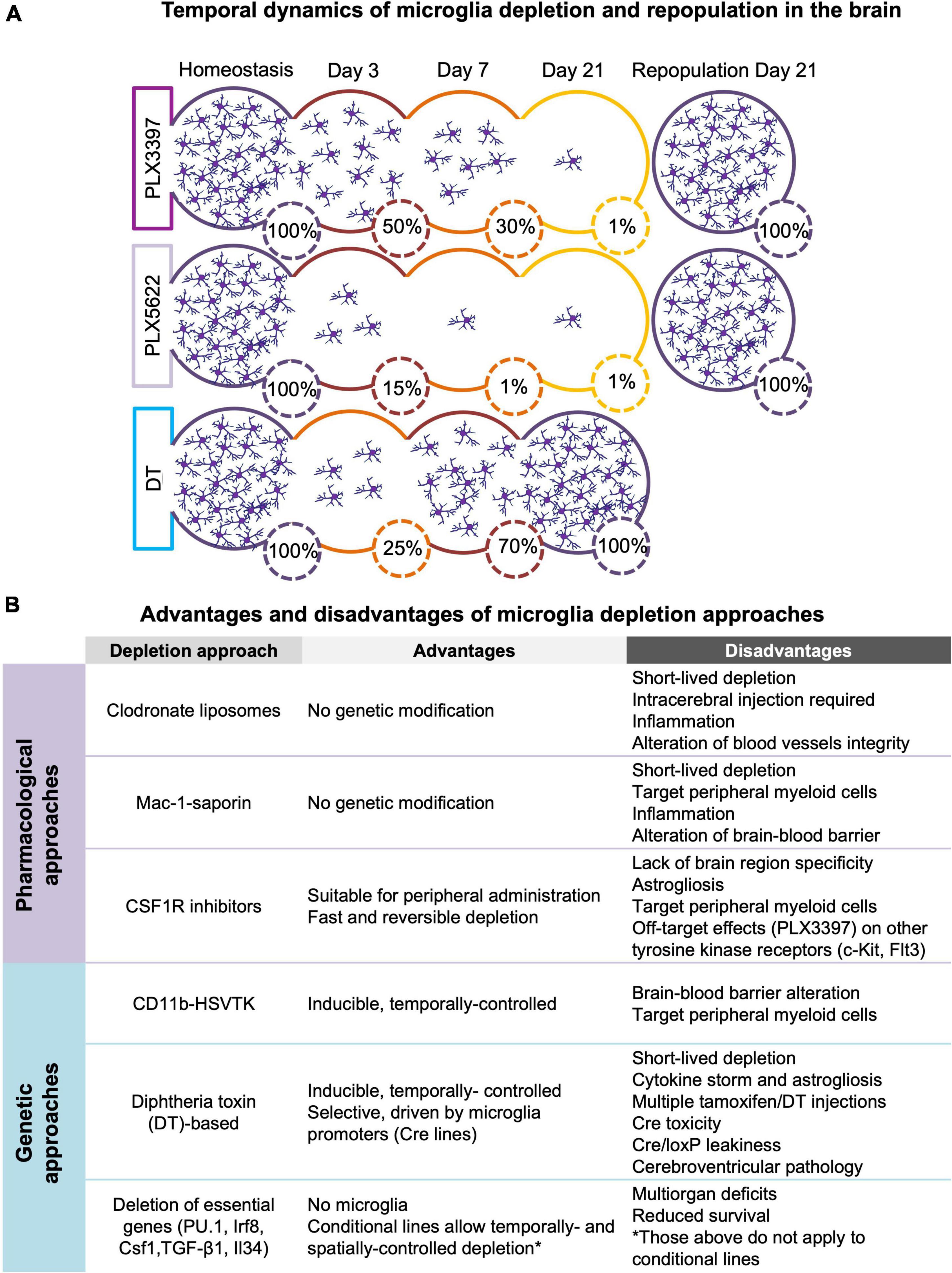
Models of microglia depletionThe possibility of acute depletion of microglia in an adult animal has contributed to highlight the role of these cells as important mediators of normal brain functioning. Here, we summarized the genetic (section 2.1) and pharmacological (section 2.2) approaches used for microglia depletion in vivo along with their respective advantages and technical limitations. Furthermore, we briefly described the impact of different depletion approaches on other central nervous system (CNS)-associated macrophages (section 2.3). Finally, we discussed the process of microglia repopulation after their depletion (section 2.4) and briefly introduced new chemo-genetic approaches to manipulate microglia function (section 2.5) Refer to Figure 1 for a comprehensive overview of the section.
Figure 1. (A) Temporal dynamics of microglia depletion and repopulation. Time-course of microglia depletion during administration of CSF1R inhibitors (PLX3397 and PLX5622) and repopulation upon treatment withdrawal, and time-course of genetic microglia depletion and repopulation induced by diphtheria toxin (DT) administration. Represented percentages of microglia in the brain have been extrapolated from published data (Elmore et al., 2014, 2018; Bruttger et al., 2015; Spangenberg et al., 2019; Basilico et al., 2021). (B) Advantages and disadvantages of microglia depletion methods. Summary of the main advantages and disadvantages of the different approaches used to deplete microglia (see section 2 for more detail).
Genetic approaches for microglia depletionThe genetic approaches for microglia depletion are based on two strategies: the expression of ‘suicidal genes’ under the control of specific microglial promoters (section 2.1.1) and the genetic manipulation of the factors essential for microglia survival (section 2.1.2).
Expression of ‘suicidal genes’ (herpes simplex virus thymidine kinase or diphtheria toxin receptor)The first approach used for depleting microglia genetically was the generation of CD11b-HSVTK transgenic mice (Heppner et al., 2005). These mice express the suicide herpes simplex virus thymidine kinase (HSV-1 TK) gene under the control of the myeloid promoter Cd11b. The HSV-1 TK exerts its toxic activity after the treatment with antiviral drugs like ganciclovir, which are further processed into toxic triphosphates by cell-intrinsic kinases resulting in apoptosis (Heppner et al., 2005). A main limitation of this approach is that the systemic and chronic administration of these antivirals can disturb the peripheral myeloid population. To avoid this limitation, these drugs must be delivered via intracerebroventricular injections (Bennett and Brody, 2014).
An alternative approach consists in the use of diphtheria toxin (DT)-based transgenic mice (Parkhurst et al., 2013; Bruttger et al., 2015). Mice carrying the DT or its receptor diphtheria toxin receptor (DTR), downstream of loxP-flanked STOP sequences, are crossed with myeloid promoter-driven Cre recombinase lines, exploiting the exclusive microglia expression of CX3CL1 receptor (CX3CR1) in the CNS (i.e., Cx3cr1Cre, Cx3cr1CreER) (Sauer and Henderson, 1988; Nagy, 2000). Microglia are then depleted upon DT administration, which inhibits intracellular protein synthesis leading to acute cell death (Bruttger et al., 2015). The use of inducible lines such as the Cx3cr1CreER has the advantage of temporally controlling both the Cre-mediated recombination and the cell ablation with DT, allowing to specifically target microglia over the other myeloid populations (Parkhurst et al., 2013). The limitations of the DT/DTR models are the astrocyte activation and the increase in cytokine production, likely released by reactive astrocytes (Bruttger et al., 2015). Other limitations include their dependence on Cre recombinase which might be toxic (Schmidt et al., 2000; Schmidt-Supprian and Rajewsky, 2007) and the use of tamoxifen (for inducible Cre lines) that binds to macrophages and other estrogen receptor-expressing cells (Donocoff et al., 2020; Green et al., 2020). Recently, some studies raised additional concerns regarding the inducible genetic models targeting microglia. The first is the potential ‘leakiness’ of the Cre/LoxP system into other brain cells or the occurrence of recombination in microglia in absence of tamoxifen induction (Chappell-Maor et al., 2020). Because leakiness varies among the mouse lines created to target microglia, the proper characterization of these Cre lines is necessary (Zhao et al., 2019; Stifter and Greter, 2020). Another concern is the cerebroventricular pathology observed in some of the transgenic lines that seems to be inherent to the ROSA-26iDTR (iDTR) allele after DT treatment, independently of Cre expression (Bedolla et al., 2022). The interpretation of the data obtained using these transgenic lines should be taken with caution. For instance, neuronal cell death (Rubino et al., 2018) and synapse degeneration (Wang et al., 2016) have been reported in these mouse lines, but not with pharmacological depletion approaches. Further validation of these genetic models is needed to improve the confidence of the findings deriving from their application.
Deletion of essential factors for microglia maturation and survivalThis genetic strategy relies on transgenic mice carrying null mutations in genes critical for microglia development such as the myeloid-specific transcription factor PU.1, interferon-regulatory factor 8 (Irf8), colony stimulating factor 1 (Csf1) and transforming growth factor beta 1 (TGF-β1). However, these constitutive models are not suitable for long-term studies as the lack of these essential genes in myeloid cells leads to reduced survival and multiorgan deficits (Shull et al., 1992; Ginhoux et al., 2010; Butovsky et al., 2013; Kierdorf et al., 2013).
To overcome this limitation, Badimon et al. (2020) recently developed a model of inducible ablation of the interleukin-34 (Il34) specifically in neuronal progenitor cells (NestinCre) that resulted in the loss of neuron-associated microglia in striatal gray matter and a model of inducible ablation of Csf1 that led to the loss of microglia in the striatal white matter. This method can also be used to target microglia subpopulations surrounding specific neuronal subtypes by ablating Il34 under different neuronal promoters (i.e., D1 and D2 receptor-expressing neurons).
Pharmacological strategies for microglia depletionPharmacological strategies for microglia depletion are also available and have potential for the clinical practice. The main pharmacological strategies are treatments with toxic-clodronate (section 2.2.1), antibodies against Mac-1-saporin (section 2.2.2) and CSFR1 inhibitors (section 2.2.3).
Toxic-clodronate-containing liposomesThe clodronate-containing liposomes are a toxin-based method to deplete macrophages and microglia that takes advantage of cellular phagocytosis (Van Rooijen and Sanders, 1996). Liposomes, small artificial vesicles, are normally phagocyted by microglia (and macrophages) leading to clodronate accumulation and ultimately resulting in apoptosis (Van Rooijen et al., 1997). However, since liposomes do not cross the bran-blood-barrier (BBB) they must be infused directly into the brain region of interest (Andreou et al., 2017; Han et al., 2019). The main limitation of this approach is that it may induce an inflammatory state and alteration of the blood vessels integrity (Han et al., 2019).
Mac-1-saporinThe immunotoxin Mac-1-saporin is an antibody against the macrophage antigen complex-1 (Mac-1), a β2-integrin glycoprotein expressed on the cell surface of most myeloid cells including microglia, conjugated to the ribosome-inactivating protein saporin (Dommergues et al., 2003; Montero et al., 2009). Intracerebroventricular or intrathecal Mac-1-saporin injection induces a transient microglia depletion followed by a rapid repopulation in rodents (Heldmann et al., 2011; Yao et al., 2016). The main limitations of this approach rely on several side effects like inflammatory reactions and BBB breakdown (Yao et al., 2016).
Inhibition of colony stimulating factor 1 receptor signaling pathwayA popular pharmacological depletion strategy consists in the inhibition of the colony stimulating factor 1 receptor (CSFR1), essential for macrophages and microglia proliferation, differentiation and survival (Stanley et al., 1997). Few inhibitors have been developed so far: PLX3397, PLX647 (Elmore et al., 2014), the more selective compounds PLX5622 (Spangenberg et al., 2019) and Ki20227 (Xie et al., 2020; Jiang et al., 2022). The most used compounds, PLX3397 and PLX5622 can cross the BBB and are usually formulated in the rodent chow for chronic administration. Many studies demonstrated a relatively fast (over 3 days) and efficient reversible depletion of microglia in the CNS (Elmore et al., 2014; Spangenberg et al., 2019; Green et al., 2020).
The main limitation of using CSF1R inhibitors (CSF1R-i) through diet is the lack of a brain region specificity. A second limitation of CSF1R-i are their putative peripheral effects (Green et al., 2020; Han et al., 2020; Lei et al., 2020, 2021; Green and Hume, 2021) since CSF1Rs are expressed on other myeloid cells (Stanley et al., 1997). Moreover, PLX3397 also binds other tyrosine kinase receptors (i.e., c-Kit and Flt3) and, at high doses, it decreases circulating monocytes in both humans and rodents (Tap et al., 2015; Butowski et al., 2016; Shi Y. et al., 2019). Notably, recent studies showed that PLX5622 reduces a population of circulating Ly6Clow monocytes responsible for the surveillance of the blood vessels (Otxoa-de-Amezaga et al., 2019) and affects peripheral myeloid and lymphoid cell populations, causing irreversible long-term changes in the bone marrow, spleen and blood (Lei et al., 2020). Furthermore, in the context of viral encephalitis, the reduction of neuroinflammation after PLX5622 treatment is not only dependent on microglia depletion but also on the reduced proliferation of mature monocytes (Ly6Chigh) in the bone marrow (Spiteri et al., 2022). On the other hand, some studies did not observe any effects in circulating monocytes and tissue macrophages when PLX3397 or PLX5622 was used at the doses typically formulated in the rodent chow (Elmore et al., 2014; Spangenberg et al., 2019; Green et al., 2020).
It must be noticed that CSF1R-i treatment has been reported to affect other brain cells. Indeed, treatment with CSF1R-i decreases oligodendrocyte precursor cells (OPC) gene expression and number, without producing major effects in the number of mature oligodendrocytes and myelination (Hagemeyer et al., 2017; Green et al., 2018; Janova et al., 2018; Garcia-Agudo et al., 2019; Liu et al., 2019). Furthermore, CSF1R-i increase mRNA and protein levels of astrocytic markers (i.e., GFAP) (Elmore et al., 2015; Basilico et al., 2021), with no changes in cell number or morphology (Elmore et al., 2014; Ma et al., 2020; Liu et al., 2021). Notably, GFAP alterations are maintained upon microglia repopulation when treatment is discontinued (Basilico et al., 2021). Recently, it has been reported that CSF1R-i treatment weakens astrocyte gap junctional coupling (Du et al., 2022). Importantly, alterations in astrocytes have also been reported with genetic microglia depletion models (Bruttger et al., 2015; De Luca et al., 2020).
While CSF1R inhibition does not induce gross changes in brain volume, neuronal gene expression (Elmore et al., 2014), neuronal density (Elmore et al., 2018), and intrinsic neuronal membrane properties (Liu et al., 2021), it may cause the accumulation of apoptotic cells in brain regions with high neuronal turnover such as the cerebellum, the olfactory bulb and the dentate gyrus (Ayata et al., 2018).
Despite all the aforementioned caveats, CSF1R-i have been widely employed for microglia depletion due to their ease-of-use in preclinical studies.
Effect of microglia depletion strategies on other central nervous system-associated macrophagesCentral nervous system immune surveillance is not only exerted by microglia but also by other CNS-associated macrophages located in the meninges, the perivascular space, and the choroid plexus that modulate the immune response at the CNS borders. As mentioned above (sections 2.2.1 and 2.2.2), the clodronate-containing liposomes and Mac-1-saporin strategies were developed to target both microglia and macrophages, rendering these tools not specific solely for the microglia population. CNS-associated macrophages, like microglia, are dependent on the CSF1/CSF1R axis for their survival and proliferation and might be affected by CSF1R-i. Specifically, a sustained treatment (21 days) with PLX3397 fully depletes meningeal and choroidal CNS-associated macrophages in healthy mice (Van Hove et al., 2019). In the context of pathology, in a murine model of viral encephalitis, treatment with PLX5622 also depletes a subpopulation of CNS-associated macrophages (Spiteri et al., 2022). Moreover, microglia and CNS-associated macrophages also share other myeloid markers such as CD11b, Iba-1, and CX3CR1, whose promoters have been used for genetic microglia targeting, and therefore commonly used genetic microglia depletion strategies fail to distinguish between microglia and CNS-associated macrophages (Goldmann et al., 2016; Van Hove et al., 2019; Prinz et al., 2021).
Microglia repopulationMicroglia cells display an extraordinary capacity of regeneration and repopulation in the brain tissue even after depletion protocols able to deplete microglia up to 90% (Elmore et al., 2014; Huang et al., 2018; Zhan et al., 2019). Although the origin of repopulating microglia is still matter of debate (section 2.4.1), acute microglia depletion and subsequent repopulation have been suggested as a therapeutic strategy to re-establish homeostasis in the diseased brain (section 2.4.2).
The origin of repopulating microgliaIn the context of pharmacological depletion strategies, Elmore et al. (2014) first proposed that repopulation (3 weeks PLX3397 + 3 weeks withdrawal) occurred from brain Nestin+ neuroectodermal progenitors, de facto excluding the proliferation from surviving or infiltrating cells as primary source for the new-born microglia. However, this hypothesis has been questioned on two grounds: (1) the origin of microglia from the yolk sac during development and (2) lack of evidence supporting the presence of microglia progenitors in the adult brain (Ginhoux and Prinz, 2015). Huang et al. (2018) were the first to refute the Nestin+ progenitor hypothesis demonstrating, through fate-mapping approach that the repopulation depends on the proliferation of residual microglia. The authors did not observe any fluorescently labeled microglia in mice with permanently labeled Nestin+ cells and their progeny (Nestin-CreERT2:Ai14 mice) after treatment (2 weeks PLX5622 + 2 weeks of withdrawal). These findings speak against a possible Nestin+ progenitor. On the contrary, they observed that repopulation occurred by proliferation of the surviving microglia cells in mice expressing fluorescently labeled microglia (Cx3cr1-CreER:Ai14 mice). Importantly, Weber et al. (2019) obtained similar results after treatment (2 weeks PLX5622 + 3 weeks of withdrawal) in Cx3cr1CreER–YFP/+R26tdTOM mice.
Even in the context of genetic microglia depletion, the origin of microglia repopulation is still under debate. Furthermore, recent evidence suggests that it is also largely dependent on the strategy being used. By using Cx3cr1CreER/+R26iDTR/+ mice, Bruttger et al. (2015) observed a robust repopulation sustained by the self-maintenance of tissue-resident macrophage populations without contribution of external progenitors. Conversely, Lund et al. (2018) observed a competitive repopulation process leading to a mosaic of cells derived from both proliferating microglia and brain infiltrating Ly6Chi monocytes in Cx3cr1CreER–YFP/+R26DTA/+ mice. The monocyte-derived cells retained transcriptional and functional differences from resident microglia, being more proinflammatory even after long-term integration into the CNS. More recently, Zhou et al. (2022) identified a subpopulation of microglia exhibiting high CX3CR1 expression and devoid of the genetic modification in Cx3cr1CreER–YFP/+R26DTA/+ mice. This subpopulation, less than 1% of all the microglia under homeostatic conditions, eluded the depletion and proliferated extensively, representing a big portion of the repopulated microglia pool.
To our knowledge, little is known about the molecular signaling that modulates and drives microglia repopulation in the brain. Bruttger et al. (2015) hypothesized the involvement of the interleukin-1 receptor signaling, showing that the administration of an IL1R antagonist impaired repopulation by affecting the proliferation of microglia clusters. Another potential candidate is the CX3CL1/CX3CR1 pathway, whose signaling exerts a positive effect on microglial repopulation (Zhan et al., 2019; Zhou et al., 2022).
The therapeutic potential of repopulating microgliaMicroglia repopulation after an acute depletion has been proposed as a promising therapeutic strategy to ‘reset’ microglia to a homeostatic phenotype in brain disease conditions (Cartier et al., 2014; Butowski et al., 2016).
In the healthy adult brain repopulated microglia after PLX3397 withdrawal display similar inflammatory transcriptomic profiles and responsiveness to pro-inflammatory stimuli to resident microglia (Elmore et al., 2015). On the other hand, microglia repopulation in the aged brain ‘reverses’ age-related microglia morphological changes, alterations in neuronal gene expression and deficits in hippocampal long-term potentiation, resulting in an improvement of cognitive functions (Elmore et al., 2018). Similarly, after brain injury microglia repopulation promotes brain recovery (Rice et al., 2017) and improves cognition (Henry et al., 2020; Willis et al., 2020). It has also been suggested that homeostatic genetic signatures displayed by the repopulated microglia might prevent its switching to a disease-associated microglia phenotype, essential to limit Aβ plaque–mediated tau pathogenesis in Alzheimer’s disease (AD) (Casali et al., 2020; Gratuze et al., 2021).
However, in some models of brain pathology microglia repopulation has been found to be ineffective (stress; Lehmann et al., 2019; Weber et al., 2019) or even detrimental [autoimmune encephalitis (EAE)]; (Rubino et al., 2018). The effects of microglia repopulation on behavioral and cognitive functions in brain disease models are described in section 4.3.
Microglia manipulation through a chemogenetic approachRecently, a new strategy based on the Gq- and Gi-coupled Designer Receptor Exclusively activated by a Designer Drug-(DREADD) technology has been developed which enables the selective stimulation or inhibition of microglia (Armbruster et al., 2007; Roth, 2016).
The DREADD technology is based on the control of engineered G protein-coupled receptors-mediated signaling. Upon activation, these receptors undergo structural rearrangements facilitating the interaction with a family of canonical heterotrimeric G-proteins (e.g., Gq, Gs, Gi, and G12/13) leading to activation of intracellular signaling pathways. The engineered receptors can be activated by the pharmacologically inert compound clozapine-N-oxide (CNO) which leads to different modulations of microglia function based on the type of G-protein associated to them (Armbruster et al., 2007; Zhu et al., 2016).
Recently, DREADDs developed under the CD68 (Grace et al., 2016, 2018) and Cx3cr1 (Saika et al., 2020; Klawonn et al., 2021; Yi et al., 2021) microglia/macrophage promoters have allowed for the manipulation of microglia. Most of these studies applied the DREADD technology to manipulate microglia function in the context of pain research. The authors found that microglia activation via Gq DREADDs induce allodynia (see Box 1) (Grace et al., 2018; Saika et al., 2020), whereas microglia inhibition via Gi DREADDs attenuate neuropathic pain and associated neuroinflammation (Yi et al., 2021). Recently, Bolton et al. (2022) also used a Gq-DREADDS under the Cx3cr1 promoter to activate microglia in a model of early life adversity and found that this manipulation normalized microglia process dynamics, synapse density and adult hormonal and behavioral stress responses. Notably, the mechanisms and effects of Gi- or Gq-DREADD on microglia intracellular signaling are still poorly understood (Grace et al., 2018; Saika et al., 2020).
BOX 1 Behavior definitions and behavioral procedure descriptions.
Spontaneous locomotor activity is typically assessed to exclude gross sensorimotor effects of pharmacological compounds or genetic alterations. It is typically evaluated in unconditioned response tests based on the exposure of rodents to novel environments. These tests also evaluate “anxiety-like” behavioral responses of rodents to unfamiliar aversive places. The open field test (OF), elevated-plus maze (EPM), and light/dark tests are based on ethological behavior that harnesses the conflict between the innate exploratory behavior of rodents in response to mild stressors (i.e., novel environments) and their aversion to open, elevated and brightly illuminated spaces (Belzung and Griebel, 2001; Bourin and Hascoët, 2003; Walf and Frye, 2007; Seibenhener and Wooten, 2015).
Repetitive behaviors: innate behaviors displayed by rodents, such as marble burying and self-grooming, might become repetitive/perseverative, like observed in rodent models of psychiatric illness or neurodevelopmental disorders. Repetitive behavior might be sensitive to anxiolytic and antidepressant drugs (Thomas et al., 2009; Ahmari, 2016).
Sensory perception, which includes vision, audition, olfaction, gustation and somatosensation, informs the brain about the environment and influences how the animal responds to it (Burn, 2008).
Nociceptive tests measure the latency or the threshold of the paw or tail withdrawal reflex after the application of electrical, thermal, mechanical, or chemical noxious stimuli. Mechanical allodynia is the response to a mechanical non-nociceptive stimulus, which is typically incremented in rodent models of chronic pain, while thermal hyperalgesia refers to an increased pain sensitivity due to an altered temperature perception (Barrot, 2012; Inoue and Tsuda, 2018). Nociception differs from pain perception, the latter including sensory, cognitive, and emotional aspects associated with damage (Barrot, 2012).
Sensory-motor tests, such as Gait Test, Grip Strength, Beam Walk, Bar Test, Hurdle Test and Pole Test, are typically used to assess gross motor deficits in mice after stroke or injury (Balkaya et al., 2013). The Rotarod Test is widely used to evaluate balance and motor coordination in mice. Standard protocols use a maximum acceleration of 40 or 60 r.p.m., with schedules ranging from 3 to 9 trials over 1 to 3 days. Performance in the rotarod improves with training allowing the evaluation of motor skill learning. Parkhurst et al. (2013) used a protocol consisting of an acceleration from 0 to 100 r.p.m. and a schedule of 60 trials/day over 3 consecutive days that allowed them to evaluate motor learning (Shiotsuki et al., 2010; Parkhurst et al., 2013).
Spontaneous alternation in the Y- or T-shaped maze can be interpreted as a measure of spatial working memory. When allowed to freely explore the apparatus, mice with an intact cognitive capacity typically remember the previously visited arms and show a tendency to enter a less visited arm (Kraeuter et al., 2019).
Sociability, the innate tendency of an animal to engage in social investigation is typically assessed in the Three-Chamber Test (also known as Crawley’s test) in rodents. It measures the preference to investigate a conspecific (located under a wire cup in one of the compartments) vs. an inanimate object. A lack of preference indicates a deficit in pro-social behavior or social interaction. In a subsequent phase of the test, the propensity of rodents to investigate a novel conspecific more thoroughly than the previous encountered one (familiar) is measured. A lack of preference for the novel mouse indicates either social novelty avoidance or poor social recognition memory. C57BL/6 or 6J mice (typical experimental strains) also display sociability toward mice of other strains (e.g., CD-1 mice) (Crawley, 2007; van der Kooij and Sandi, 2012; Ayash et al., 2020).
Recognition learning and memory, the ability to discriminate a previously encountered item as familiar, is typically evaluated with the Novel Object Recognition (NOR). The test exploits the fact that mice display an innate tendency to spend more time investigating a never encountered (novel) object than a familiar one. A variant of this task is the Novel Object Location (NOL), in which only the location of the object in the arena is changed between the familiarization and test phases. A third variant is the novel Context-Object-Discrimination (COD) in which is measured the capacity to discriminate the congruency between the objects and the context where they were previously encountered. The performance of these tasks depends on the integrity of hippocampus, perirhinal, and medial prefrontal cortices circuitry (Broadbent et al., 2009b; Barker and Warburton, 2015).
Fear conditioning is a type of associative learning achieved by a Pavlovian/Classical conditioning procedure in which a neutral conditioned stimulus (CS), typically an auditory cue, is associated to an aversive unconditioned stimulus (US), often a mild electric foot-shock. In presence of the CS rodents express freezing behavior, a species-specific fear response, indicative of the memory of the auditory cue with the aversive stimulus (Auditory or Cued Fear Conditioning, AFC). Animals also exhibit fear responses when returned to the chamber in which the tone and shock were paired, or a chamber in which shocks occur alone (Contextual Fear Conditioning, CxFC). The hippocampus is required for the formation and retrieval of context–fear associations, whereas the amygdala is required for conditioning and recall of associations to both contextual and discrete cues (LeDoux, 2003).
Spatial learning refers to the process by which an animal makes a mental representation of the surrounding environment that permits its orientation and location of relevant places. Among the most used paradigms are the Morris Water Maze (MWM) and the Barnes Maze (BM). Both tests use the innate motivation of the animal to find a safe location, a submerged platform in the case of the MWM, and a hidden hole in the case of the dry BM. The animal must rely in distal cues to learn to navigate the maze and find the target. These tests evaluate hippocampally dependent spatial learning and reference memory (Vorhees and Williams, 2006; Gawel et al., 2019).
‘Depressive-like’ behaviors in rodents typically refer to symptoms of helplessness, anhedonia, and behavioral despair since the full spectrum of depressive symptoms cannot be recapitulated in animal models. The expression of despair can be evaluated by measuring immobility in the Forced Swim Test (FST) and Tail Suspension Test (TST), which place the animal in inescapable distressful situations. The learned helplessness model measures the rate of non-escape responses displayed by rodents following an uncontrollable and inescapable stress when re-exposed to the same situation. A decrease in sucrose preference is a measure of anhedonia in rodents. All these measures are sensitive to the action of antidepressants (Krishnan and Nestler, 2011).
Effects of microglia depletion on synaptic function and plasticityOver the last decades, increasing evidence revealed that microglia are involved in the regulation of synaptic plasticity under physiological conditions. Microglia contact axonal boutons and dendritic shafts in an activity-dependent manner resulting in the elimination and/or formation of spines and pre-synaptic elements (Wake et al., 2009; Tremblay et al., 2010; Paolicelli et al., 2011; Schafer et al., 2012; Hayashi et al., 2013; Lim et al., 2013; Miyamoto et al., 2016; Sipe et al., 2016; Weinhard et al., 2018; Basilico et al., 2019; Wang and Huang, 2020). In this section, we reported the effects of microglia depletion on structural plasticity (section 3.1), neurogenesis (section 3.2) and functional changes in synaptic transmission (section 3.3) in the adult brain. Refer to Figure 2 for a comprehensive overview of the section.
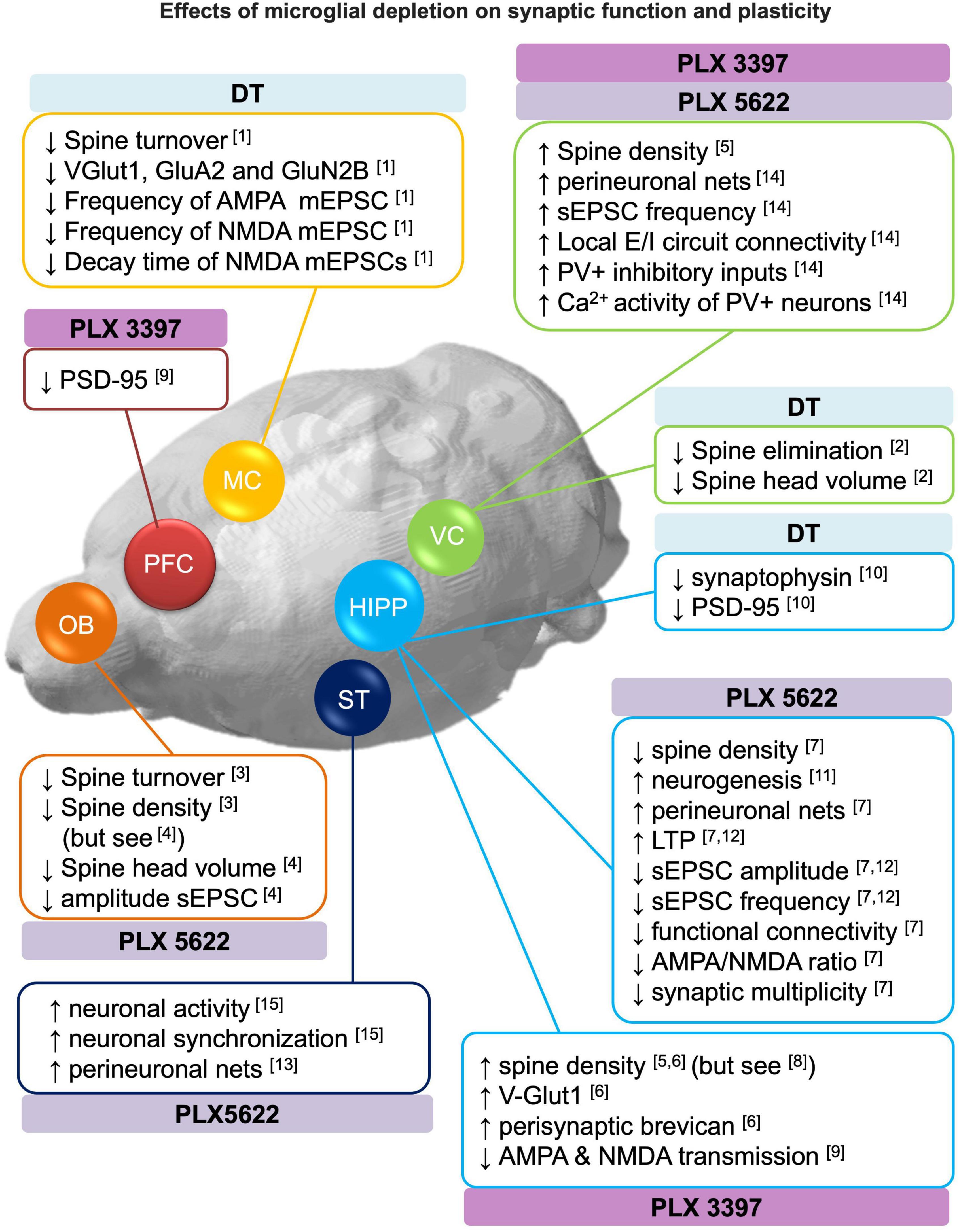
Figure 2. Effects of microglial depletion on synaptic function and plasticity. Schematic representation of the reported effects of microglia depletion with CSF1R inhibitors (PLX3397 and PLX5622) and genetic DT-induced on dendritic spines, neurogenesis, perineuronal nets, excitatory and inhibitory (E/I) neurotransmission (see section 3 for more detail). DT: diphtheria toxin, HIPP: hippocampus, MC: motor cortex, OB: olfactory bulbs, PFC: prefrontal cortex, ST: striatum, VC: visual cortex. [1] Parkhurst et al., 2013; [2] Ma et al., 2020; [3] Reshef et al., 2014; [4] Wallace et al., 2020; [5] Rice et al., 2015; [6] Strackeljan et al., 2021; [7] Basilico et al., 2021; [8] Pinto et al., 2020; [9] Yegla et al., 2021; [10] De Luca et al., 2020; [11] Elmore et al., 2018; [12] Corsi et al., 2022; [13] Crapser et al., 2020; [14] Liu et al., 2021; [15] Badimon et al., 2020.
Structural plasticitySynaptic contacts are structurally plastic, and connectivity is remodeled through mechanisms of synapse formation, stabilization and elimination. Although structural plasticity is maximal during brain development, in the adult brain it is important for learning and memory and to restore function after brain damage (Xu et al., 2009; Caroni et al., 2012). In this section, we reported the effects of microglia depletion on the formation, elimination and/or stabilization of dendritic spines, and the effects of microglia depletion in perineuronal nets.
Perineuronal nets (PNN) are components of the extracellular matrix that modulate local inhibitory transmission and plasticity in the adult brain. PNNs have been related to the stabilization of synaptic connections, playing a role in synaptic maturation and the formation and consolidation of memories (Gogolla et al., 2009). Alterations in PNNs are also present in diseases where memory deficits are core features (Carulli and Verhaagen, 2021). Pharmacological microglia depletion has been reported to increase density and intensity of perineuronal nets in several brain regions (Crapser et al., 2020; Basilico et al., 2021; Liu et al., 2021) and to increase the expression of peri-synaptic extracellular matrix proteoglycan brevican (Strackeljan et al., 2021). These results are in line with recent evidence reporting that microglia engulf components of the extracellular matrix in response to neuronal cytokine signaling (Nguyen et al., 2020). The functional implications of the role of microglia on degrading PNN might vary depending on age, brain-region and or inflammatory conditions. For instance, Tansley et al. (2022) showed that after nerve injury, degradation of PNNs by microglia enhances the activity of projection neurons contributing to pain-related behaviors.
Concerning the dynamics of dendritic spines, three studies on microglia depletion (both genetic and pharmacological) consistently reported a reduced spine turnover in the motor cortex (Parkhurst et al., 2013), visual cortex (Ma et al., 2020) and granule cells (abGC) of the olfactory bulb (Reshef et al., 2014) as revealed by in vivo two-photon imaging.
On the other hand, the analysis of spine density after microglia depletion provided heterogeneous results. In particular, spine densities were increased in: visual cortex [DT genetically induced microglia depletion (Ma et al., 2020); PLX3397 (4 weeks) (Rice et al., 2015)], olfactory bulb-abGCs [PLX5622 (4 weeks) (Wallace et al., 2020)] and hippocampal CA1 [PLX3397 (4 weeks) (Rice et al., 2015; Strackeljan et al., 2021)]. Spine densities were decreased in: olfactory bulb abGCs [PLX5622 (4 weeks) (Reshef et al., 2014)] and hippocampal CA1 [PLX5622 (1 week) (Basilico et al., 2021)]. No effect was observed in the hippocampus of juvenile mice (P22) [PLX3397 (1 week) (Pinto et al., 2020)]. These conflicting findings outlined above can be recapitulated in the context of spine turnover. Indeed, it has been suggested that in homeostatic conditions spine turnover is very dynamic and rarely impacts long-term network structure (Berry and Nedivi, 2017). It is therefore plausible that the net spine density is an insufficient measure to capture the effects of microglia depletion on spines. Furthermore, the method or duration of microglia depletion may also contribute to the variability observed on spine density measures.
Spine morphology may be indicative of the stabilization and maturation of spines, correlates with synaptic strength and has been implicated in memory consolidation processes (Berry and Nedivi, 2017). Some studies suggest that microglia depletion diminish the rate of mature spines, revealed by an overall reduction in spine head size (Ma et al., 2020; Wallace et al., 2020), an increase of immature thin or filopodia-like spines (Rice et al., 2015; Wallace et al., 2020) or a reduction in mature spines (mushroom, stubby, and bifurcated) (Rice et al., 2015; De Luca et al., 2020; Ma et al., 2020). On the other hand, three studies reported no effects of microglia depletion on spine morphology (Reshef et al., 2014; Basilico et al., 2021; Strackeljan et al., 2021). As discussed above the discrepant results are likely due to methodological differences between studies.
The impact of microglia on dendritic spine turnover and density is not easy to define, since the dynamics of persistent and transient spines have different impacts on the local circuit (Berry and Nedivi, 2017), and, as discussed above for PNNs, it might depend on age, brain-region and or inflammatory conditions. Despite this, two studies highlighted that the role of microglia on spine regulation might be relevant for learning and memory and behavioral performance. In this regard, Parkhurst et al. (2013) observed that reduced spine turnover in the motor cortex was associated with a reduced motor learning rate in mice with depleted microglia. Furthermore, Wang C. et al. (2020) observed that microglia depletion increased spine density in hippocampal engram cells whose reactivation correlated with the recall of a contextual fear memory (see section 4.1.7).
Altogether, these data suggest that microglia actively participate in circuit remodeling by modulating spine turnover and PNN degradation.
NeurogenesisNeurogenesis, a process of generation of new neurons, represents another form of brain structural plasticity which occurs in two regions of the adult brain, i.e., olfactory bulb and the dentate gyrus of the hippocampus, by integrating newly born neurons into already existing circuits (Van Praag et al., 2002; Lledo et al., 2006; Rodríguez-Iglesias et al., 2019).
It has been reported that microglia depletion with PLX5622 increased the number of dividing neurons [bromodeoxyuridine (BrDU+)] in the dentate gyrus (Elmore et al., 2018). Notably, Wang C. et al. (2020) reported that microglia depletion (PLX3397 5 weeks) prevented memory forgetting induced by the pro-neurogenic drug memantine, suggesting a role of microglia in hippocampal neurogenesis-related plasticity (see section 4.1.7). However, no effects have been reported on the number of new-born cells [doublecortin (DCX+)] in the olfactory bulb (Reshef et al., 2017). These apparent conflicting findings are in line with Reshef et al. (2014) who reported that microglia-mediated fractalkine signaling was involved in hippocampal- but not olfactory bulb-related neurogenesis, suggesting that the modulation of neurogenesis by microglia is regional specific. Diaz-Aparicio et al. (2020) also showed that microglia balance proliferation and survival in the hippocampal neurogenic niche by secreting neurogenic modulatory factors following the phagocytosis of apoptotic cells.
In a model of traumatic brain injury (TBI), where survival of immature neurons (DCX+) was compromised, microglia depletion (genetic DT-induced and PLX5622) further decreased immature neuron survival. However, upon repopulation microglia promoted immature neuron survival (Willis et al., 2020).
Altogether, the evidence suggest that microglia in the healthy brain positively regulate hippocampal neurogenesis and related plasticity and that its repopulation after injury promote functional recovery by supporting immature neuron survival.
Functional plasticityFew in vivo imaging studies demonstrated that microglia monitor and respond to neuronal activity (Wake et al., 2009; Tremblay et al., 2010; Pfeiffer et al., 2016). Recent evidence also suggest that microglia can suppress neuronal activity (Badimon et al., 2020) or regulate neuronal synchronization (Akiyoshi et al., 2018). These findings suggest a neuron-microglia bidirectional communication that promotes functional changes in synapses and neuronal circuits. In this section, we summarized the main findings observed on the regulation of excitatory (section 3.3.1) and inhibitory (section 3.3.2) neurotransmission after microglia depletion.
Excitatory neurotransmissionStudies employing microglia depletion strategies indicate that microglia regulate the level of synaptic proteins and the functioning of glutamatergic excitatory synapses, likely in a brain region-dependent manner.
In the motor cortex, Parkhurst et al. (2013) observed a decrease in the abundance of the synaptic proteins (i.e., GluN2B, VGlut1, and GluA2) after genetic DT-induced microglia depletion in juvenile mice. These changes were associated with reduced frequencies of miniature excitatory postsynaptic currents (mEPSCs) in layer V pyramidal neurons, suggesting a decrease in spontaneous glutamate release. Moreover, the decay time of NMDA mEPSCs was reduced, consistent with the observed reduction of GluN2B subunits. These synaptic alterations were associated with deficits in learning-induced dendritic spine remodeling and decreased performance in learning tasks dependent on microglia BDNF signaling (refer to section 4).
In the hippocampus, three studies showed that the absence of microglia leads to decreased excitatory synaptic transmission (Basilico et al., 2021; Yegla et al., 2021; Corsi et al., 2022). In particular, Basilico et al. (2021) demonstrated that microglia depletion with PLX5622 (1 week) led to increased long-term potentiation (LTP), reduced glutamatergic transmission and reduced functional connectivity at CA3-CA1 hippocampal synapses. These changes were associated with immature synaptic features such as reduced AMPA/NMDA ratio and impaired multiplicity. Similarly, Corsi et al. (2022) observed increased plasticity and reduced excitatory neurotransmission following PLX5622 (1 week). Both studies highlighted the importance of the CX3CR1/CX3CL1 axis in microglial control of synaptic function (Basilico et al., 2021; Corsi et al., 2022). A third study showed that treatment with PLX3397 (3 weeks) was associated with decreased synaptic transmission in both young and aged rats. In particular, through extracellular field recordings in hippocampus, the authors showed an impairment in both AMPAR- and NMDAR-mediated transmission at CA3-CA1 synapses (Yegla et al., 2021). During microglia repopulation after genetic DT-induced depletion, a reduction of synaptophysin and PSD-95 (but not Vglut1, Glun2A,2B) was observed in the hilus of the hippocampus (De Luca et al., 2020). Decreased amplitude, but not frequency of sEPSC was also observed in abGC neurons of the olfactory bulb devoid of microglia (PLX5622 4 weeks), suggesting weaker excitatory inputs (Wallace et al., 2020).
On the other hand, in layer V pyramidal neurons of the visual cortex, microglia depletion with PLX3397 and PLX5622 (3–4 weeks) in adult mice enhanced spontaneous EPSC (sEPSC) frequency, but not amplitude, suggesting an increase in presynaptic release. The authors also observed an increase in both excitatory and inhibitory synaptic connections to excitatory cortical neurons assessed with a functional circuit mapping experiment (Liu et al., 2021). Similarly, Badimon et al. (2020) showed that microglia depletion with PLX5622 (3 weeks) resulted in increased neuronal activity and synchronization in the mouse striatum revealed by in vivo two-photon calcium imaging, suggesting that microglia suppress neuronal activation in this brain circuit. The authors also identified a negative feedback mechanism involving the purinergic signaling. This consisted in the ability of microglia to sense ATP released upon neuronal activation through the purinergic receptor P2RY12. Subsequently, microglia convert ATP in AMP and then adenosine, which negatively modulates activity in striatal D1-expressing neurons acting on adenosine A1AR receptors.
Altogether, these data point to a brain region-dependent role of microglia in regulating excitatory synaptic function. In support of this regional disparity, recent studies reported a phenotypic heterogeneity of microglia between brain regions (Grabert et al., 2016; De Biase et al., 2017; Ayata et al., 2018). Notably, the microglia depletion approaches used in the aforementioned studies (i.e., DT-induced and PLX) lack regional specificity and hence deplete microglia in the whole brain. Consequently, compensatory mechanisms occurring between brain areas cannot be ruled out.
Inhibitory neurotransmissionLittle is known about the impact of microglia depletion on GABAergic inhibitory neurotransmission. The only study showing GABAergic alterations (Liu et al., 2021) reported that microglia depletion (PLX3397 3 weeks) upregulated parvalbumin (PV) expression and calcium activity of inhibitory PV+ interneurons in the visual cortex, suggesting increased PV+ inhibitory inputs onto L5 pyramidal neurons in the microglia-depleted brains. On the other hand, (Ma et al., 2020) did not observe changes in miniature inhibitory postsynaptic currents (mIPSC) in the L2/3 pyramidal neurons of the visual cortex in juvenile mice treated with PLX3397 (2 weeks). Likewise, microglia depletion with PLX5622 did not induce changes in spontaneous inhibitory postsynaptic currents (sIPSC) in the adult hippocampus (Basilico et al., 2021) or abGC neurons of the olfactory bulb (Wallace et al., 2020).
Like the evidence regarding glutamatergic neurotransmission (section 3.2.1), these data point to a microglia brain region-dependent regulation of inhibitory synaptic function. Likely, microglia exert a fine control of the excitation-inhibition balance depending on local micro-environmental cues.
Effects of microglia depletion on behaviorFor decades, the study of behavior, learning and memory has focused only on neuronal mechanisms. Activity-dependent synaptic plasticity has been proposed to be the substrate of learning and memory (Bliss and Lømo, 1973; Takeuchi et al., 2014; Sweatt, 2016; Humeau and Choquet, 2019). Recent evidence suggested that microglia respond to neuronal activity and modulate synaptic transmission, plasticity and behavior (Yirmiya and Goshen, 2011; Morris et al., 2013; Parkhurst et al., 2013; Tay et al., 2018; Rodríguez-Iglesias et al., 2019; Badimon et al., 2020; Wang C. et al., 2020; Cornell et al., 2022).
In this section, we summarized the recent literature on the impact of microglia depletion on behavior in the healthy adult animal (section 4.1), impact of microglia depletion during development on adult behavior (section 4.2) and microglia depletion and behavior in rodent models of disease (section 4.3).
Effects of microglia depletion during adulthood on behavior in healthy rodentsIn this section, we reviewed the effect of microglia depletion on spontaneous locomotor activity (section 4.1.1), sensory perception (section 4.1.2), motor and performance learning (section 4.1.3), sociability and social recognition memory (section 4.1.4), recognition learning and memory (section 4.1.5), associative learning and memory (section 4.1.6), and spatial learning and memory (section 4.1.7). Refer to Figure 3 for a comprehensive overview of the section.
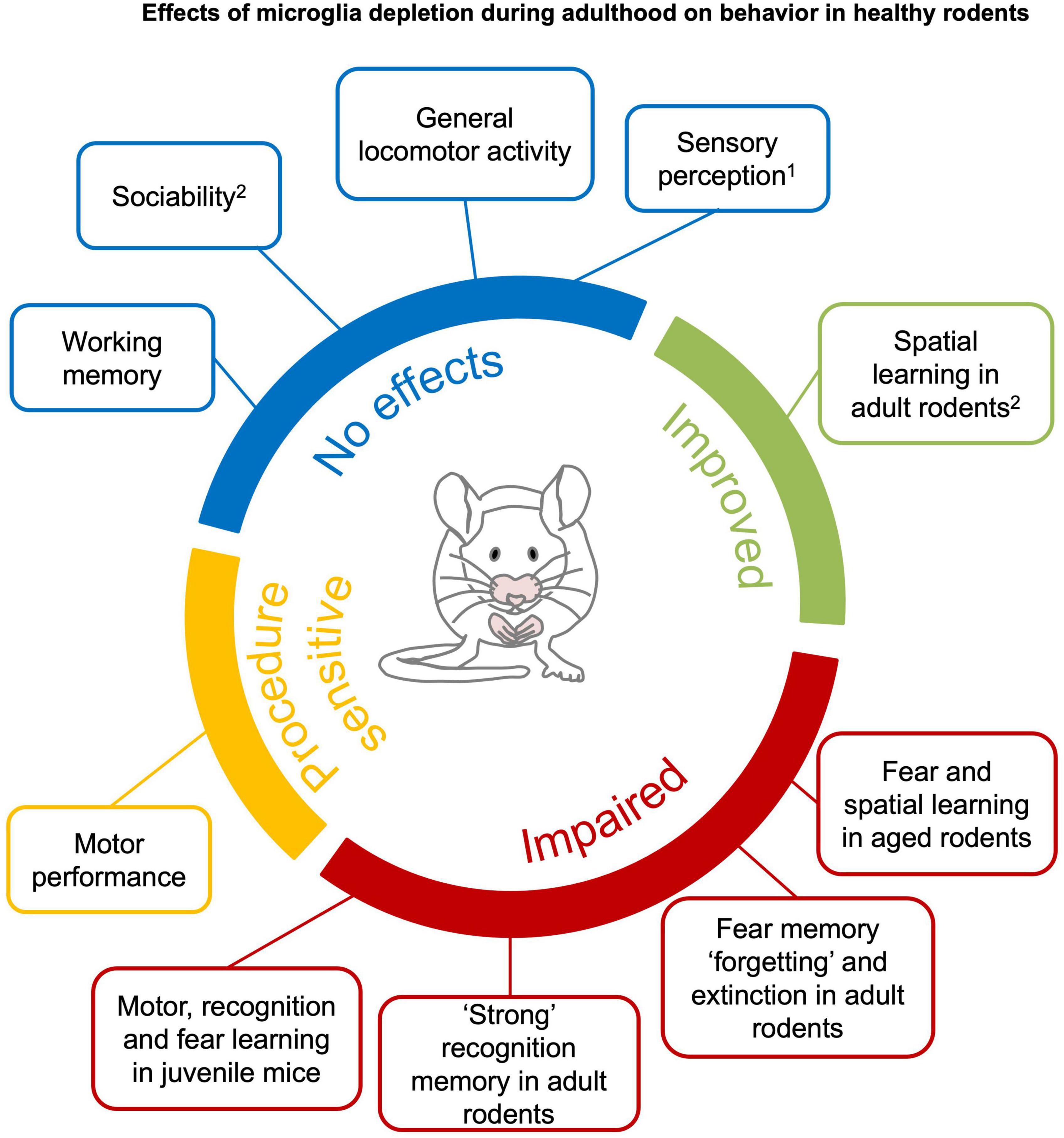
Figure 3. Effects of microglia depletion during adulthood on behavior in healthy rodents. Microglia depletion in the healthy adult brain does not have major effects on sensory perception1, general locomotor activity, anxiety, sociability and working memory (blue). Motor function is impaired after genetic DT-induced microglia depletion (Parkhurst et al., 2014; Rubino et al., 2018), but not pharmacological depletion (yellow). Complex recognition memory and specific learning processes, such as “forgetting” and extinction of fear memories, are compromised in young mice with microglia depletion. Learning deficits are observed when microglia are depleted in juvenile or aged mice (red). Spatial learning is improved in young mice (green).1An exception is reported by Rubino et al. (2018) that found altered gustatory perception in genetic DT-induced microglia depletion. 2Clodronate injection in the hippocampus have been reported to impair sociability and spatial learning in young mice (Torres et al., 2016). See section 4.1 and Supplementary Tables 1–8 for detailed information.
Spontaneous locomotor activitySpontaneous locomotor activity in a novel environment is typically assessed in preclinical experimental settings to exclude gross sensorimotor effects of pharmacological compounds or genetic alterations (Seibenhener and Wooten, 2015). In general, the literature consistently showed that depletion of microglia does not affect locomotor activity, ‘anxious-like’ or repetitive behaviors (see Box 1), independent of the species (mice or rats), age, method and duration of microglia depletion (Supplementary Table 1) (Elmore et al., 2014, 2018; Dagher et al., 2015; Rice et al., 2015; Torres et al., 2016; Ayata et al., 2018; Adeluyi et al., 2019; De Luca et al., 2019; Lehmann et al., 2019; Spangenberg et al., 2019; Weber et al., 2019; Allen et al., 2020; Badimon et al., 2020; Wang C. et al., 2020; Wu et al., 2020; Xie et al., 2020; Basilico et al., 2021; Cao et al., 2021). Similarly, no effects on locomotor activity have been reported upon microglia repopulation (up to 2–3 weeks after depletion with PLX3397 or clodronate liposomes) (Elmore et al., 2014; Torres et al., 2016). There are two noticeable exceptions, Vichaya et al. (2020) who observed reduced voluntary wheel running in absence of microglia (PLX5622 4 weeks) and Elmore et al. (2018) who observed increased exploration of a novel environment after 4 weeks of microglia repopulation.
Sensory perceptionDeficits in sensory perception (Box 1) may affect behavioral performance. For this reason, it is critical to exclude any sensory deficit caused by microglia depletion. At present, there are a few studies that addressed sensory competence in microglia depletion models. Particularly, they investigated visual, gustatory, nociceptive and olfactory perception (Supplementary Table 2).
Wang et al. (2016) evaluated whether sustained genetic tamoxifen-induced microglia depletion (4 weeks) influenced the structure and function of the retina in mice. The authors found that visual acuity—measured by elicited motor reflexes in response to light stimuli—was intact, despite they observed degeneration of photoreceptor synapses. Two studies reported that microglia depletion with PLX does not affect nociception (Box 1) in absence of previous lesion or damage (Liang et al., 2019; Sawicki et al., 2019). Badimon et al. (2020) reported that microglia depletion (PLX5622, 3 weeks) did not affect performance in a task that relies on olfaction in mice, suggesting that this sensory modality is intact in absence of microglia.
On the other hand, De Luca et al. (2019) observed that genetic DT-induced microglia depletion in rats reduced food and water intake. By employing an extensive battery of behavioral and neurobiological assays, the authors concluded that the absence of microglia altered the hypothalamic-thalamic gustatory circuitry, resulting in a strong distaste for palatable foods (i.e., saccharin, sucrose, high-fat, and high-sugar diets).
Given the paucity of studies explicitly addressing gustatory perception, to gain a hint on this matter, we reported the results from other studies that monitored food and fluid intake in the absence of microglia. Consistent with the aforementioned study, pointing to an alteration of the gustatory system, chronic microglia depletion in mice with PLX5622 (8 weeks) reduced the consumption of high-fat diet but not standard diet (Valdearcos et al., 2017). On the contrary, shorter PLX3397/PLX5622 treatment (1–5 weeks) did not affect mice consumption of standard diet, high-fat diet, sucrose or alcohol (Gao et al., 2019; Ali et al., 2020; Cai et al., 2020; Wang J. et al., 2020; Cansell et al., 2021; Warden et al., 2021). The available evidence do not allow to draw a clear conclusion and future studies should address these discrepancies, likely arising from diverse microglia depletion methods and durations. Future studies also should take into account that microglia depletion with PLX may alter microbiota composition (Yang et al., 2021) and metabolic parameters (Ali et al., 2020).
Motor performance and motor learningRegarding motor performance (Box 1), genetic but not pharmacological microglia depletion has been associated with motor impairment (Supplementary Table 3).
Pharmacological microglia depletion with CSFR1 inhibitors in mice did not induce general motor deficits in a variety of sensory-motor tests (Qu et al., 2017; Yang et al., 2018; Garcia-Agudo et al., 2019; Crapser et al., 2020; Henry et al., 2020; Li et al., 2020; Xie et al., 2020) and it did not induce alterations on balance and motor coordination as revealed by the rotarod test (Elmore et al., 2014; Qu et al., 2017; Yang et al., 2018; Liang et al., 2019; Badimon et al., 2020; Crapser et al., 2020). Similarly, no effects were observed upon microglia repopulation (Elmore et al., 2015; Henry et al., 2020).
On the contrary, using genetic DT-induced microglia depletion, Parkhurst et al. (2013) observed that juvenile mice displayed reduced rate of motor learning improvement which was associated with a reduced motor learning-induced spine turnover. By using a similar conditional animal model, Rubino et al. (2018) reported that microglia depletion and subsequent repopulation triggered ataxia and motor incoordination. The authors also observed neuronal death in the somatosensory cortex and an inflammatory signature in repopulated microglia.
These conflicting results suggest that the effects of microglia depletion on motor function might be sensitive to the approach used for microglia depletion.
Sociability and social recognition memorySociability and social recognition memory (see Box 1) are not affected by pharmacological whole-brain microglia depletion (Supplementary Table 4). In particular, mice treated with either PLX3397 or PLX5622 did not display deficits in social preference and recognition memory of conspecifics (Torres et al., 2016; Badimon et al., 2020) nor deficits in the interaction with mice of other strains (Lehmann et al., 2019). Subsequent microglia repopulation did not affect those social behaviors either (Lehmann et al., 2019; Gu et al., 2021).
On the contrary, microglia depletion with clodronate injections within the hippocampus reduced social preference and recognition, a deficit that recovered upon microglia repopulation. Notably, in the same study, mice treated with PLX3397 (3 weeks) did not display social deficits (Torres et al., 2016). A possible explanation for this discrepancy are the inflammatory side effects produced by the intracerebral injection of clodronate (Han et al., 2019), since an inflammatory state might cause reduced social exploration and social withdrawal (Moieni and Eisenberger, 2018).
留言 (0)