
記住我







In a previous study, we tested the response of 100 domestic cats to N. cataria, A. polygama, L. tatarica, and V. officinalis [1]. Results from that study indicated that cats who did not respond to N. cataria (catnip) often responded to at least one of the other three plants. Because plants were available to the cats for up to only 1 h, we limited our analysis to scoring the absence or presence of the “catnip response” and did not study their behavior in detail. Here we studied the response of 6 domestic cats (Table 1) in their familiar, permanent home environment to the same 4 plants used in our previous study, plus Acalypha indica (Indian nettle) (Fig. 1, Table 2, Additional file 1: Figure S1A-F), which has not been tested before to our knowledge. To allow for a comprehensive analysis of cat behavior in response to the cat-attracting plants, each plant was presented to the cats, spread over 10 days (Additional file 1: Figure S2A). This dataset was analyzed for differences in (1) response duration and (2) behavior in response to these plants between (A) the cats and (B) the plants tested.
Table 1 Age, breed, hair-color and pattern, and gender of the cats who participated in the studyFig. 1
Botanical illustration of the cat-attracting plants used in this study. Not all tissues of these plants have an effect on cats. Cats respond to (from left to right) the barkless wood of Lonicera tatarica, the leaves of Nepeta cataria, the roots of Acalypha indica and Valeriana officinalis, and to the woody stem, leaves, and kiwi fruit galls of Actinidia polygama. The silver vine fruit galls (deformed fruits) are induced by the female gall midge Pseudasphondylia matatabi, who lays her eggs in the A. polygama flower buds. The illustration of P. matatabi was based on photographs of Pseudasphondylia kiwiphila and Pseudasphondylia elaeocarpi. This botanical illustration was created by Vicky Earle from Draw in Nature Studio and was commissioned by Cowboy Cat Ranch
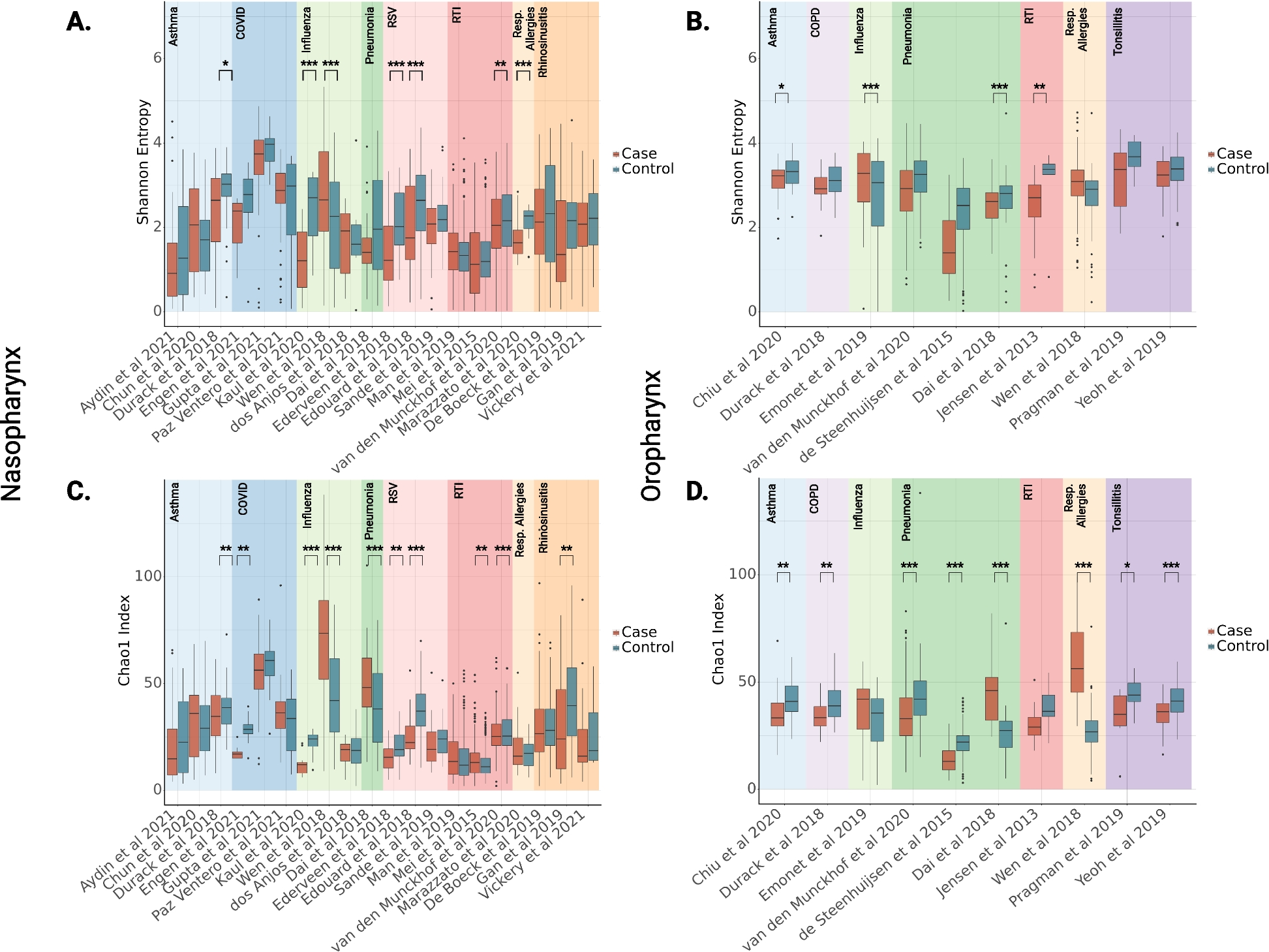
Table 2 An overview of the plant materials that were used in this studyAll but one of the 6 cats responded to all 5 plants tested (Fig. 2 and Additional file 1: Figure S3), and all responses to the plants could be classified as “catnip responses”, meaning the cats showed (a combination of) behaviors listed in Table 3 and shown in Additional file 2: Video 1. We observed approximately 2 h of responses to A. polygama and L. tatarica, 1.5 h to N. cataria and A. indica, and 1 h to V. officinalis. Since 5 of the 6 cats in this study had never responded to N. cataria in the past, two different brands of catnip were used to investigate whether fluctuations in the level of active compounds in different sources of catnip could account for variation in (or lack of) attractiveness. One sock contained catnip from the brand Frontier, the other from the brand SmartyKat. When comparing the daily total response duration to both catnip brands for each cat separately, we observed that cat O responded significantly longer to the catnip from Frontier (Additional file 1: Figure S4). This finding suggests there may be a difference between the two brands of catnip that were used in this study, but overall, many and robust responses were observed from all 6 cats to catnip from both brands.
Fig. 2
Response duration and response frequency of domestic cats to cat-attracting plants. Box and whisker plots showing the total response time, median response duration, and the total number of responses of 6 domestic cats to 5 cat-attracting plants. Each dot represents the data of one cat; the middle line in the bars shows the median value. Each cat-attracting plant was available for 100 h, the control (green tea) was available for 500 h (100 h for each of the 5 plants tested). A Data shown per plant. Note the large spread of the data points, indicating large variation in response duration and frequency to the various plants between the cats. Differences between the 5 plants (total response duration, median response duration, and response frequency) were not statistically significant (P > 0.05, mixed-effects repeated measures ANOVA and Tukey post hoc test, corrected for multiple comparisons). We obtained 5 instead of 6 data points for V. officinalis since cat H was unable to participate due to medical reasons. For the statistical analysis of the paired data with missing data (cat H), we used a parametric test (mixed-effects repeated measures ANOVA). Therefore, for the analysis we used the average values (both the average response time to a plant for each cat and the average of the cats for each plant) instead of the median. Using either the average or median data did not affect the outcome of the statistical analysis. B Differences in total response time, median response duration, and response frequency between cats. Colors represent the fur color of the cats. Response duration and frequency differed significantly between the cats (Kruskal-Wallis). P values shown in the graph are from Dunn’s post hoc tests. *P < 0.05; **P < 0.01, ***P < 0.001
Table 3 Ethogram describing body positions and behaviors seen in domestic cats in response to cat-attracting plants or their volatile compoundsWhile previous work had suggested domestic cats respond euphorically to A. indica (Indian nettle) root in a similar fashion to catnip [18], this plant has never been tested on cats in a controlled study. Since the cat-attracting effect of A. indica root quickly disappears after harvest [18] and its geographical distribution does not extend to North America, roots were lyophilized immediately after collection on Christmas Island, Australia, in an attempt to preserve their effect on cats. Our data show that the response duration to the lyophilized roots of Indian nettle was similar to the other plants that were tested.
The cats only sparsely interacted with the negative controls (green tea). The total response time (any engagement with the object, not behavior specific to the “catnip response”) from all cats to the negative controls after 500 h availability was just over 6 min, which is approximately 1% of the observed response time to the cat-attracting plant materials (490 min). Nearly all interactions with the negative control were from cat V and most of them occurred when A. polygama was tested. Three cats never engaged with the negative controls.
There was no statistically significant difference in total response time of the cats between the 5 plants (Fig. 2A). Total response time is the sum of the duration of all responses and is determined by both response frequency and response duration. We also did not find a statistically significant difference in the median response duration and response frequency of the cats between the cat-attracting plants. However, when comparing the response duration to the 5 different plants between the 6 cats, we found these to be significantly different (Fig. 2B). Cats O and N responded longer to the cat-attracting plants than cat Z. The differences in total response time to the cat-attracting plants between the cats could be explained by both differences in the length of the responses and the frequency of responses. These data show there are significant differences between cats in how long and frequently they respond to cat-attracting plants.
There was no statistically significant difference in response duration between the various plants, possibly because of the large variation between the cats. However, when we looked at the responses to the various plants for each cat individually, we observed that cat H responded significantly longer to A. polygama and cat O to L. tatarica and N. cataria than to some of the other plants (Fig. 3). Interestingly, cat Z showed no interaction at all with the sock containing V. officinalis root over the full 5-week testing period.
Fig. 3
Response duration to cat-attracting plants shown for each cat individually. Each dot represents the total response duration of 1 day (10 h), with the middle line in each box showing the median of these 10 days. Each plant was available for 10 days (total of 100 h). Note that the Y axes are not the same for all graphs since the goal was to illustrate differences between the plants for each cat, not between cats. The Kruskal-Wallis test was used to test for statistically significant differences between plants. P values shown in the graph are from Dunn’s post hoc tests. *P < 0.05; ***P < 0.001
The data also show that N. cataria (catnip) was not more popular than the other plants tested when comparing across the 6 domestic cats in this study. The longest total response duration after 100 h, as well as the longest total response per day, and the longest single response was never to N. cataria (Additional file 1: Figure S5). These results suggest that while catnip might be the best-known cat-attracting plant among cat caregivers outside of East Asia, the other plants seem to be at least as potent.
Behavior observed for cats O and V in response to the plant Menyanthes trifoliata (buckbean) suggests this plant is also able to elicit the “catnip response”. Fifteen grams of dried buckbean leaves (Siberian Herbals) inside a sock was offered to cats A, N, O, V, and Z for a couple of hours on one day. We observed one response of cat O that lasted about half a minute and one response of cat V that lasted a little over 1 min.
The degree of attraction to cat-attracting plants differs between catsNext, we looked at the degree of attractiveness of the plants. This was measured by the time it took a cat to respond to the plant for the first time after it was made available on each of the 10 test days. The data show no difference in attractiveness between the 5 plants we tested (Fig. 4A). However, we did observe significant differences in how strongly individual cats were attracted to the plants (Fig. 4B). These results suggest that the time to first response is in part determined by the cat’s personality (consistent differences in behavior between individuals, e.g., curiosity or fear of missing out), rather than intrinsic properties of the plant. Therefore, we also compared the times to first response to the 5 cat-attracting plants for each cat separately. Seeing differences in time to first response between the plants for individual cats may suggest differences in intrinsic properties between the plants. Similar to response duration, while we did not see differences between the time to first response when we looked at the combined data of all 6 cats, we did see statistically significant differences in time to first response between plants when we analyzed the data for each cat separately (Fig. 5). While cat O did not have a single day out of the 50 without responding at least once, cat Z did not respond at all on about 70% of the days, including the 10 days V. officinalis was available. Cat O responded to L. tatarica and N. cataria almost immediately on each of the 10 test days. In contrast, the first response to V. officinalis of cat O was about 9 h on three of the 10 test days. The opposite was seen for cat V, who appeared to be attracted more strongly to V. officinalis than to N. cataria. On all 9 days that cat V responded to V. officinalis, this was within or around half an hour. These results suggest that the level of attractiveness of a plant is not solely determined by properties of the plant, but also by how the cat perceives the plant.
Fig. 4
Time to first response. A The median time till the first response of 6 cats is shown for 5 cat-attracting plants. Each dot represents the median time till the first response of 10 testing days of each cat to the cat-attracting plants. Cat H did not participate in testing V. officinalis. There were no statistically significant differences in the time to the first response between the plants (P > 0.05, mixed-effects repeated measures ANOVA (paired test with missing data; see Fig. 1A)). B The median time till the first response of 5 cat-attracting plants is shown for the 6 domestic cats. Each dot represents the median time to first response of 5 cat-attracting plants. The differences between the cats were statistically significantly different (Kruskal-Wallis). P values shown in the figure are from Dunn’s post hoc test. ** P < 0.01, *** P < 0.001
Fig. 5
Time to first response to 5 cat-attracting plants shown for each cat separately. Each dot shows the time it took the cats for their first response on each of the 10 test days. Cat H did not participate in the testing of V. officinalis. * P < 0.05 (Dunn’s post hoc test)
Taken together, these data show that all 5 plants are equally capable of attracting domestic cats and eliciting the “catnip response”, while both response duration and how strongly individual cats are attracted to the plants can differ significantly. These differences might in part be due to variation in olfactory perception and in part to differences in the cats’ personalities.
The “catnip response” is different between cats, but comparable among various cat-attracting plantsIn addition to the quantitative analysis (i.e., duration of the response), we also studied the qualitative aspects of the responses to the various plants. We created an ethogram that is specific for the “catnip response” (Table 3). Some of these behaviors may be affected by how the olfactory stimulus is offered to the cat. For example, biting and pulling with the object in the cat’s mouth will be possible when the plant material or single compound is offered inside or on a fabric, respectively, but it will not be observed when powder of dried A. polygama fruit galls is sprinkled on the floor. In this study, all plant materials and single compounds were offered on or in a fabric and therefore allowed for comparison between cats, as well as between plants or single compounds. Behaviors not mentioned and described in the ethogram either did not occur (Flehmen, lordosis, vocalization) or were not analyzed because of limitations such as camera angle and distance (e.g., drooling). Sniffing was not included because it was considered behavior used to detect or identify an odor, not behavior in response to smelling odorants. Although not specifically studied or analyzed, signs of stress, fear, or aggression (as determined by, e.g., flattened, backwards ears or a low tail close to the body) were never observed. In addition to previously described behavior in response to catnip, we have added “rippling of the back” and “twitching of the back” (Additional file 2). This behavior is not linked to feline hyperesthesia syndrome. There is no reaction (biting, scratching, or licking of the area where the twitching or rippling occurs) of the cats to the concerning area of the back, rather, the cat seems completely unaffected by it. Twitching and rippling of the back appeared to be quite specific for the “catnip response” since it was only rarely observed on other occasions. “Rolling on the side” reflects the frequency of changes in body position (standing/sitting to lying on the side or lying on the side to lying on the back). Rippling and twitching of the back, as well as rolling on the side and head shaking are extremely short events and are therefore reported and shown as events per minute response, whereas all other behaviors are reported and shown as the percentage of the total response time. The percentages can exceed 100% since some behaviors can be displayed by the cats simultaneously (e.g., holding and rubbing, or, holding and raking).
To compare behavior between the cats, we analyzed 5 responses to N. cataria nearest to 60 s of each cat using BORIS behavioral analysis software. Catnip was chosen because the variation in frequency and length of the responses of the 6 cats was least for this plant. During the response, the cats were mostly either sitting or lying on their side. Time spent while standing or lying on their back during the response was also observed, but not frequently (Fig. 6). Body position during the response varied enormously between the cats. Cat O predominantly lay on his side while engaging with the filled sock, cats A, H, and Z responded predominantly in a sitting position, and cats N and V showed an equal mix of sitting and lying on their side (Fig. 7).
Fig. 6
Heatmap showing similarities and differences in behavior between 6 domestic cats in response to N. cataria (catnip). For each cat, the five responses nearest to 60 s were analyzed using BORIS behavioral analysis software. All P values shown are from the Kruskal-Wallis test. med, median; ns, not statistically significantly different; ** P < 0.01; *** P < 0.001; **** P < 0.0001
Fig. 7
Body position and behavior of 6 domestic cats observed in response to N. cataria (catnip). Results for “biting”, “head rubbing”, “holding”, “lying on side”, and “raking” are shown as time spent relative to the total response duration (percentage), whereas results for “head shaking”, “rippling of back”, and “rolling on side” are depicted as the number of events per minute of response. Data for the body position “sitting” is not shown because sitting and lying down were mutually inclusive and inversely correlated (Fig. 5). All P values shown are from Dunn’s post hoc tests. * P < 0.05; ** P < 0.01
Our data also suggest there is large variation between cats in most behaviors that are typical for the “catnip response”. Head rubbing the olfactory object was the behavior observed most frequently, and although it was seen for all 6 cats, there were significant differences between the cats (Fig. 7, Additional file 3). The response to N. cataria for cats A and H consisted almost exclusively of head rubbing, significantly more than for cat O. In addition to head rubbing, cat O showed other behaviors such as raking or biting while holding the object. The amount of time spent holding the sock, raking, and biting was significantly greater for cat O than for several of the other cats (Fig. 7). Rippling of the back was not seen for cats A and H but was a characteristic feature of cat Z’s response, where it was seen at high frequency (Fig. 7). In fact, about 15% of her response time was rippling of the back. Head shaking, rolling on the side, and twitching of the back were seen for most or all cats, with no differences between cats for the latter. The frequency of head shaking was significantly different between the cats O and Z (Fig. 7). This behavior seemed to be rather specific for the “catnip response” since it was not seen during their normal daily activity. None of the cats had medical problems with their ears, nor did we observe any buildup of wax in their ear canal to account for head shaking. We also did not see any scratching or pawing aimed at the head or ears, which would be indicative of medical problems with the ears. Perhaps this head shaking behavior is similar to “shake-off” behavior seen in dogs where it can serve as a “reset button” after excitement, although there is no literature that would support this hypothesis. Alternatively, it might be a way for the cats to shed excess saliva, since it is known that these cat-attracting plants can induce drooling [1].
Overall, the frequency of rolling on the side was low. The responses of cats N and O seemed more dynamic than the response of cats A and Z since rolling on the side from a sternal position, or onto the back from a lateral position, was seen more frequently with cats N and O (Fig. 7). Collectively, these data demonstrate that the behavior seen in the “catnip response” is quite consistent for each cat, but show enormous variability between cats.
Having observed large variation in response traits of domestic cats towards catnip, we wondered if their idiosyncratic behavioral pattern would be the same for all the various cat-attracting plants used in this study. As can be seen in Fig. 6, the behavioral pattern in response to N. cataria is quite distinct between cats A, O, and Z. Cats A and Z have a fairly simple behavioral response where they predominantly sat and head rubbed the object, with cat Z also frequently demonstrating rippling of her back. On the contrary, cat O spent much more time lying on his side, raking, biting, and holding the object, and rolled on his side much more frequently than the other two cats. To test if there is a difference in behavioral patterns of cats towards different cat-attracting plants, we analyzed the behavior of cats A, O, and Z in response to all plants tested in this study.
During the response of cat A to any of the 5 plants, she predominantly sat and head rubbed the filled socks (Fig. 8, cat A). While some licking was seen during some of her response to A. polygama and V. officinalis, the body position and behaviors of cat A were highly similar between catnip and the 4 other plants.
Fig. 8
Body position and behavior observed during the response to various cat-attracting plants. For cats A, Z, and O five responses to each plant nearest to 60 s were analyzed using BORIS behavioral analysis software. We observed only two responses from cat Z to A. indica. Therefore, two responses instead of 5 were analyzed. P values shown are from the Kruskal-Wallis test. med, median; ns, not statistically significantly different; * P < 0.05
We observed lots of rippling of the back for cat Z in response to N. cataria. Behavioral analysis revealed that rippling of the back was not specific for catnip, but rather part of her general response since it was observed in response to all cat-attracting plants (except V. officinalis to which she never responded) (Fig. 8, cat Z). In addition to rippling of the back, we also observed twitching of the back in response to all the other plants tested. It is unknown whether rippling of the back (wavelike motion) and twitching of the back (single contraction on one location lasting a fraction of a second) are related. Her body position and behavior during the responses to the other cat-attracting plants were highly similar in proportion and frequency when compared to catnip.
Finally, we compared the behaviors of cat O between the 5 different plants. His response to N. cataria was the most diverse and complex out of all the 6 cats with him predominantly in a lateral position (~85% of the response time) when head rubbing (~50%), raking (~35%), and biting occasionally (~15%) while holding the object (~50%). Cat O rolled on his side from a sternal position 2–3 times per minute response duration, and we rarely observed headshaking (without the sock in his mouth), and rippling or twitching of his back. In line with what we observed for cats A and Z, his behavioral pattern was near identical for all cat-attracting plants (Fig. 8, cat O). The data also suggest however that holding and raking was seen less frequently for cat O when responding to V. officinalis, especially when compared to N. cataria (Fig. 8, cat O and Additional file 1: Figure S6). These findings are interesting when considering the previous observations that cat O was significantly less attracted to V. officinalis root than to N. cataria (Fig. 5) and that his total response duration to valerian root was also less than to other cat-attracting plants (Fig. 2).
Taken together, these data suggest that while responses between cats vary, the behavior of individual domestic cats to diverse cat-attracting plants is highly similar, although the effect of V. officinalis root on cats seems to be slightly different.
Response duration to cat-attracting plants decreases with repeated exposureThe setup of the experiments, with its repeated presentation, allowed us to learn more about possible habituation (reduced response duration over time to the same stimulus) to the cat-attracting plants. Information about possible habituation will be useful when giving advice to cat caregivers on how to use olfactory stimuli for environmental enrichment. Furthermore, seeing differences in habituation between plants might suggest the presence of different compounds or quantities of these compounds in the cat-attracting plants.
The olfactory stimuli were offered 2–3 days a week, for 10 h a day, for two periods of 2 weeks (weeks 1–2 and 4–5), with an interstimulus interval of at least 9 days between weeks 2 and 4 (Additional file 1: Figure S2A). First, we compared the total response time (median of 6 cats) during the first 2-week testing period (weeks 1 and 2) with the second 2-week testing period (weeks 4 and 5). When we analyzed all 5 cat-attracting plants together, we found that the median response time was the same (Fig. 9A). We observed a similar pattern when we looked at the plants individually, suggesting that either no habituation occurred within the 5-week testing period or that the 1-week interstimulus interval was sufficient to reverse any habituation that may have occurred during the first 2-week testing period.
Fig. 9
Response duration to cat-attracting plants over time. Each dot represents data (total response time) of one cat. When all plants were compared, each dot shows the median value of the total response durations to the 5 cat-attracting plants. A The total response duration of 6 cats to 5 cat-attracting plants during the first 5 testing days (50 h; weeks 1–2) was compared to the total response time during the 5 testing days (50 h) during weeks 4–5 (Additional file 1: Figure S2A). The test periods of 2 weeks were separated by a 9-day interstimulus interval. B Total daily response time of 6 cats during the first (days 1 and 6) and last day (days 5 and 10) of both 2-week testing periods. Cat H did not participate in testing V. officinalis. For all statistical analyses, the paired, non-parametric Wilcoxon matched-pairs signed rank test was used. All P values were > 0.05. Only P values < 0.1 are shown
To test the latter, we compared the response duration between day 1 and day 5, as well as between day 6 and day 10. While none of the observed differences were statistically significant, we did see a decline in response time to A. polygama within both the first and the second 2-week testing period (Fig. 9B). The response duration on the last day of both 5-day testing periods (days 5 and 10) was shorter for nearly all cats, suggesting that some habituation may have occurred. The response duration to this plant was the highest of all plants tested on the first day of both 5-day testing periods.
To learn more about possible habituation to the various stimuli, we performed additional experiments where the plant material was offered 10 days in a row for 2 or 12 h per day (Additional file 1: Figure S2B). To rule out the effects of potential degradation or complete volatilization of the active compounds over time, two new socks with fresh plant material were offered every day. Habituation was observed for A. polygama (dried fruit gall powder) and L. tatarica (sawdust) (Fig. 10, days 1–10). A similar pattern was seen for N. cataria (dried, cut leaves), but the difference between day 1 and day 10 was not statistically significant. We did not have enough material to also test A. indica. For all plants tested, after 1 to 1.5 weeks of daily, voluntary exposure (2 or 12 h a day), the response duration of each cat was reduced to (close to) zero. After the 10-day testing period and possible habituation to the plant materials, a different cat-attracting plant was offered to learn if the scent from this stimulus would result in the reappearance of the response. This dishabituation would suggest the presence of other active compounds or higher levels of similar compounds in the newly offered stimulus. After habituation of the cats to either L. tatarica, A. polygama, or N. cataria, no dishabituation was seen when the cats were offered different cat-attracting plant material (Fig. 10). The only exception was cat O, who showed a longer response to L. tatarica than his first and longest response to A. polygama and N. cataria (Fig. 10A+D), underscoring the idiosyncrasy between cats. Furthermore, these results suggest that L. tatarica may contain compounds not present, or at significantly lower amounts, in catnip and silver vine. Another interesting finding was the observation that offering N. cataria to the cats who were habituated to A. polygama and L. tatarica did not significantly increase response duration. This might suggest that nepetalactone binds to (some of) the same olfactory receptor(s) as some of the active compounds present in A. polygama and L. tatarica. These findings also indicate that offering cat-attracting plants on a non-continual basis or alternating between the various cat-attracting plants could prevent or reduce habituation in cats.
Fig. 10
Habituation and dishabituation to cat-attracting plants. The response duration of 4–5 domestic cats to three different cat-attracting plants is shown for 10 consecutive days. With habituation a gradual decrease in response duration over time is seen. Dishabituation is the reappearance or increased duration of a response to a different stimulus that is offered to the cats after habituation has occurred (day 11 or 12, or both) and its duration is similar or higher to what was seen on day 1. Results for A. polygama (A and B) are shown in red, for L. tatarica (C) in orange, and for N. cataria (D) in green. See Additional file: Figure S2B for more details. The differences between day 1 and 10 for A. polygama and L. tatarica were statistically significant (Friedman test). P values shown in the figure are from Dunn’s post hoc test. * P < 0.05; ** P < 0.01
Cat-attracting compounds in A. polygama are not exclusively produced in response to the parasitic attack of the gall midge P. matatabiBoth normal A. polygama fruit and fruit galls used in our previous study [1] were collected from vines growing in East Asia. In this natural habitat of the plant, gall midge Pseudasphondylia matatabi females can lay their eggs in the plant’s flower buds. As a result of this parasitic invasion, fruit galls develop. It seems that the presence of P. matatabi larvae in the developing kiwi fruit is critical for the synthesis of compounds that serendipitously attract cats, since we have previously shown that domestic cats respond to dried A. polygama fruit galls, but not to dried normal fruit [1]. Indeed, we were able to detect P. matatabi DNA in dried fruit galls that we used in our preceding study (Fig. 11A). Sequencings results confirmed, unequivocally, that P. matatabi DNA was present in the A. polygama fruit galls (100% percent identity and query coverage; Additional file 4).
Fig. 11
Response of domestic cats to Texas-grown A. polygama. A Detection of P. matatabi DNA in dried A. polygama fruit galls from East Asia. Species-specific primers were used to amplify a 330-bp fragment of the mitochondrial cytochrome oxidase subunit 1 gene. Sanger sequencing and nucleotide BLAST confirmed the DNA was from the gall midge P. matatabi. B Response time, shown in seconds per hour availability, of 5 cats to Texas-grown silver vine plant material. The cats were offered dried leaves from a female and male silver vine variety ("Pavel" and "Hot Pepper", respectively), as well as dried, lignified stem. The response time to dried, powdered A. polygama fruit galls originating from East Asia is shown in dark red. "Pavel" and "Hot Pepper" leaves were available to the cats for 15 and 16 h, respectively. Stem was available 2 × 15 h. Powdered silver vine galls were available for 100 h total (10 days, 10 h per day). C Observed behavior of cat Z in response to Texas-grown A. polygama stem (brown dot in panel B). Bars show either behavior expressed as the percentage of the total response time (left Y axis) or the number of events per minute response time (right Y axis; “head shaking”, “rippling of back”, and “rolling on side”). Cat Z responded 4 times to the locally grown silver vine stem, with a total response time of 74 s. Only observed behavior is shown. HP, A. polygama Hot Pepper variety; P, A. polygama Pavel variety; TX, Texas
We wondered if the gall midge induces the synthesis of these compounds only locally (fruit) or systemically (stem, leaves, fruit). It is known that some domestic cats do respond to dried A. polygama stem [1]. However, we do not know if these tissues were obtained from silver vine plants in East Asia that were bearing fruit galls at the time of harvest. Since A. polygama is dioecious and P. matatabi females deposit their eggs in the flower buds, not the fruit, one could argue that in response to oviposition in a male flower bud the plant might also systemically induce synthesis of cat-attracting compounds. However, P. matatabi oviposition in male flower buds or male flower bud galls have never been observed (Dr. Junichi Yukawa, Kyushu University, Fukuoka, Japan, personal communication, June 2021). To test whether the presence of the gall midge is required for the synthesis of the cat-attracting compounds, we grew A. polygama locally (Mico, Texas, USA), where P. matatabi does not occur. The cats were offered dried leaves from the female Hot Pepper variety and the male Pavel variety, each for almost a full day. Seeing cats respond to leaves from male plants, even when grown in their natural habitat and hence in the presence of P. matatabi, would suggest that the gall midge is not required for the production of these compounds. All five cats responded to the locally grown A. polygama leaves, both from the male and female plant (Fig. 11B). Although the data are limited, they strongly suggest the leaves were at least as popular among the domestic cats as the dried gall material from East Asia. The shorter response to the leaves from the Pavel variety may be explained by harvesting later or the longer drying time of the leaves. Harvest time for those leaves was later in the fall when the leaves would soon be shed by the plant. Testing these already collected leaves was postponed because we wanted cat A, who had recently received radioactive iodine treatment for hyperthyroidism, to also participate. Stem from the female silver vine Hot Pepper variety was made available to the cats on two different days. In agreement with our previous findings [1], only a small percentage (20%) of the cats responded to the silver vine stem. Cat Z responded 4 times: 26, 8, 18, and 22 s, with a total response time of 74 s, and analysis of her behavior showed that the response was similar to the behavior observed when exposed to the other cat-attracting plants: mostly head rubbing in a sitting position with her back rippling and an occasional head shake (Fig. 11C). No responses were seen to the control stem (lignified Juniperus ashei). Interestingly, while cat Z responded for a total time of 4 min and 15 s to the dried leaves of the Hot Pepper variety, she did not touch the sock containing the leaves for approximately half of that time. No other responses where there was no contact with the test object by cat Z or any other cat to any plant material were seen. Instead of contact with the object, she rubbed her head on the floor, rolled on her side, and her back rippled, all in close proximity (approximately 20 cm) to the olfactory object. This observed behavior in response to the dried silver vine leaves was characteristic for her and highly similar to her responses to other plants. This cat never demonstrated this behavior in response to any of the controls, which were available for hundreds of hours, and her most recent response prior to these responses was 3.5 weeks earlier. Therefore, we concluded this response was specific to the A. polygama leaves.
We previously concluded that domestic cats do not respond to A. polygama leaves grown in the USA [1]. However, subsequent DNA barcoding (matK) revealed that the leaves previously used for testing were from the closely related species Actinidia arguta instead of Actinidia polygama. These A. arguta leaves were only used for one small ex
留言 (0)