
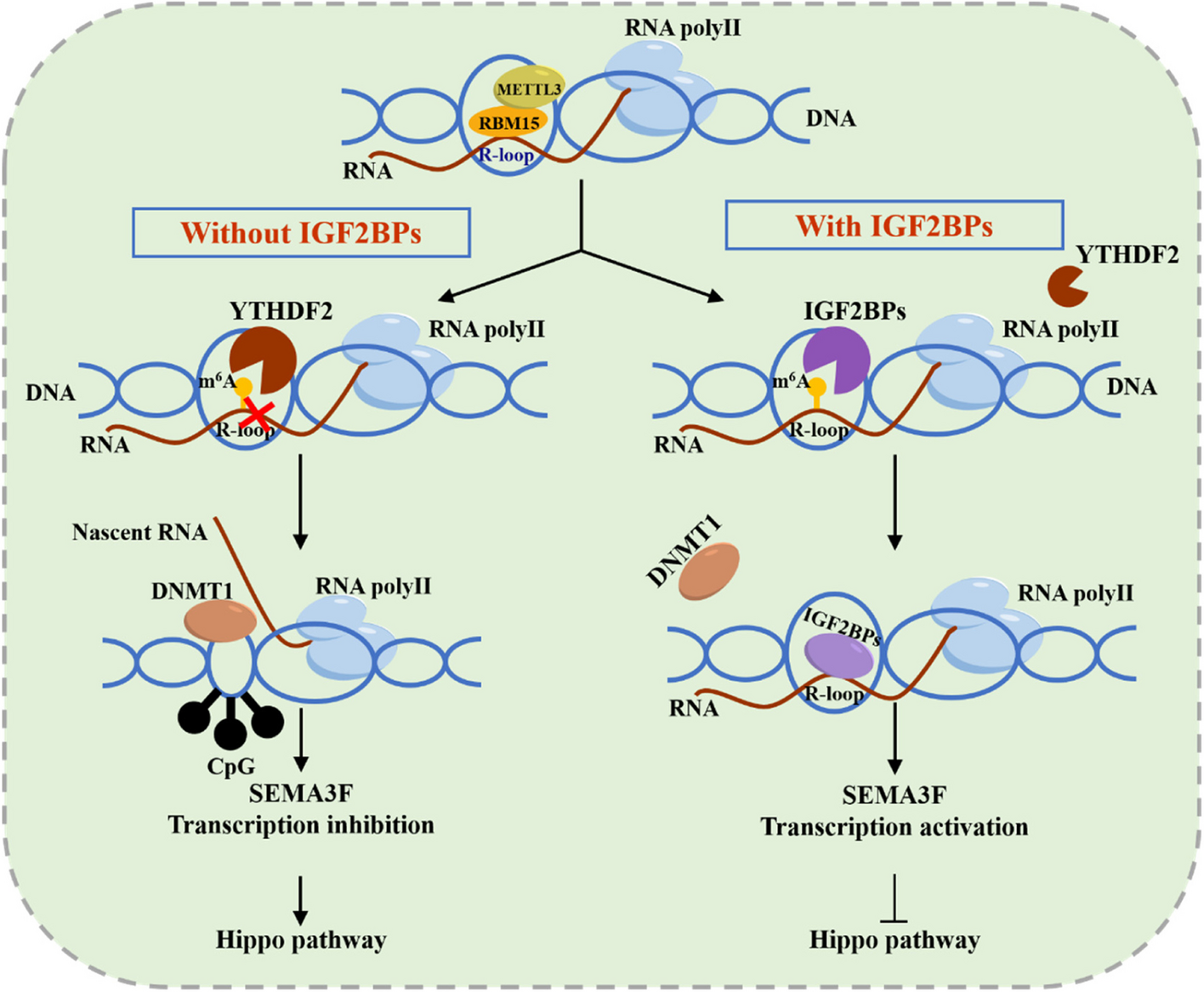
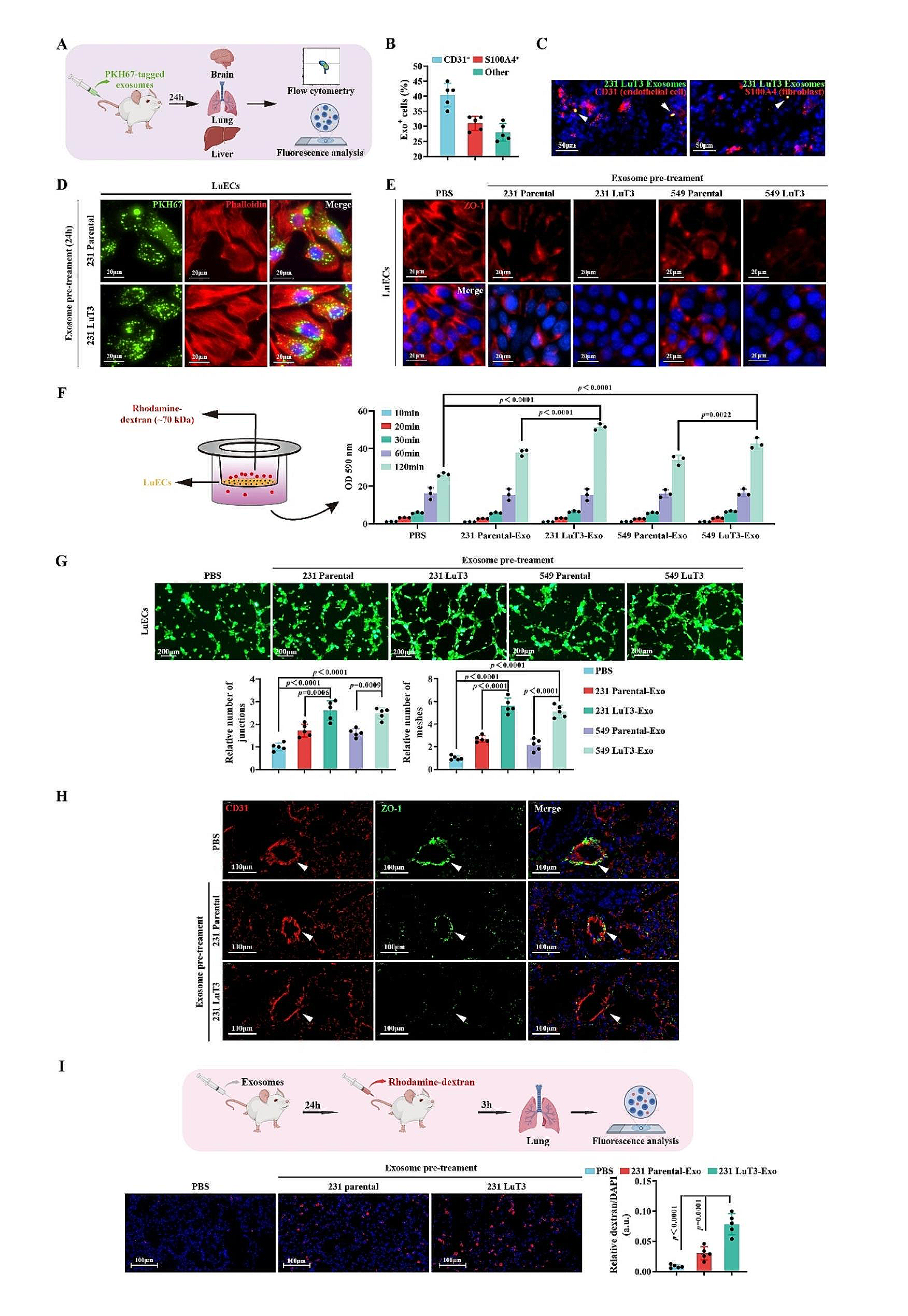
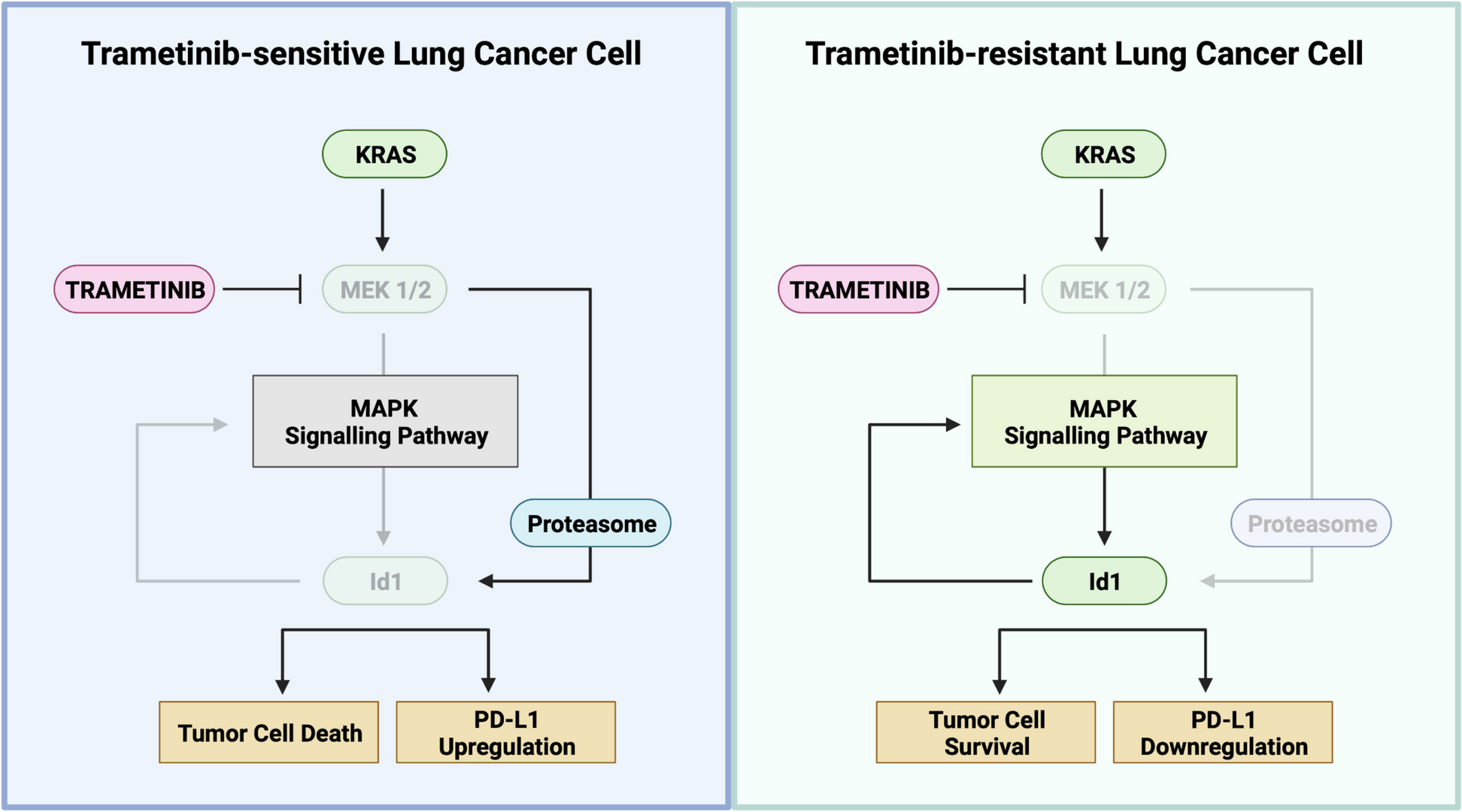
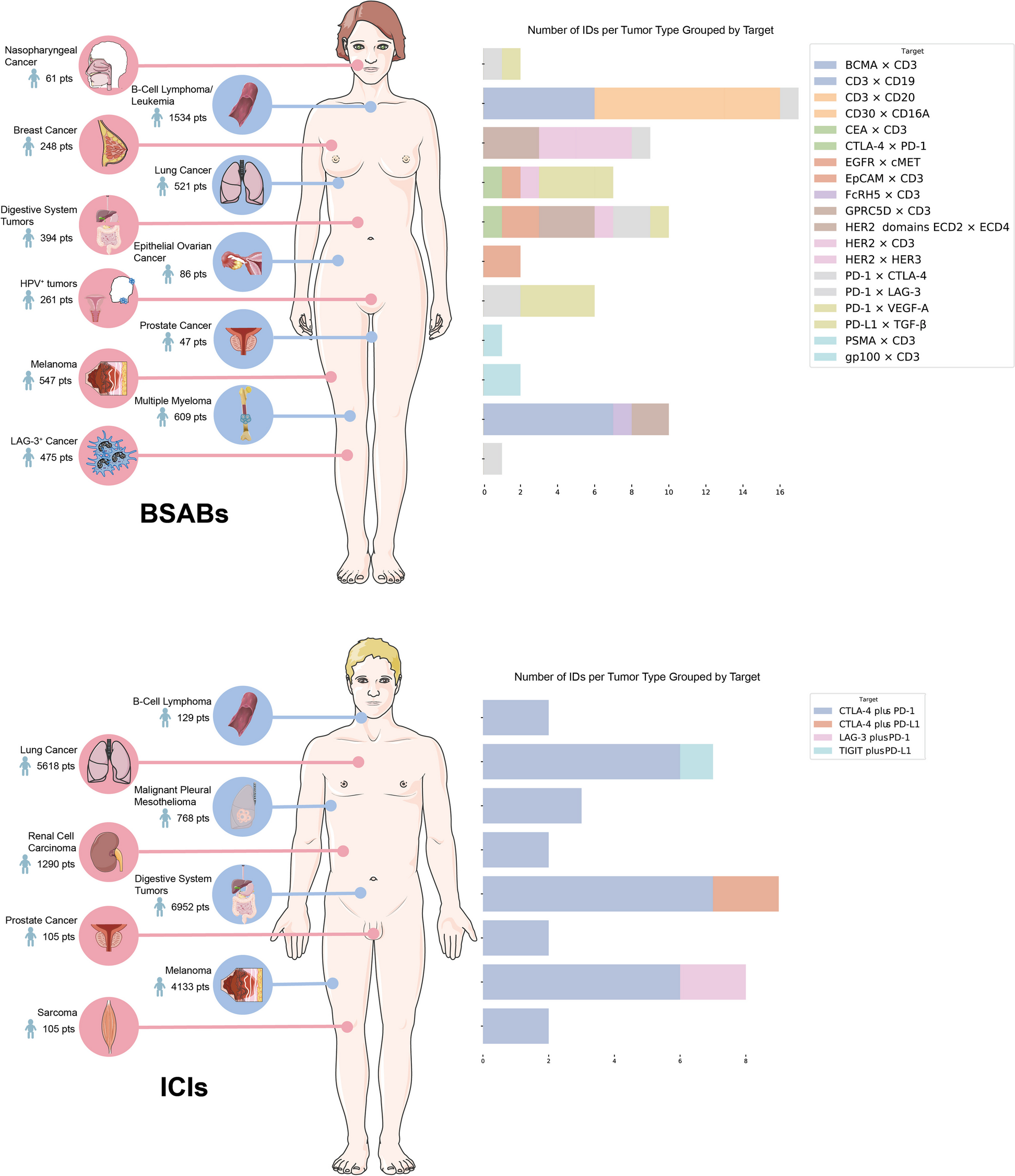
記住我


Clonal expansion and evolution is a major determinant of AML relapse [17]. To identify the genomic landscape at Dx and Re, we performed whole exome sequencing (WES) and gene fusion detection based on bulk RNA-sequencing. We detected between 4 and 26 somatic mutations per sample (Fig. 1A, Supplemental Table 2). This analysis confirmed the presence of an inframe insertion in the juxtamembrane domain (JMD) between amino acid 583 and 611 in all four FLT3-ITD patients as well as AML1-ETO fusion transcripts in the AML1-ETO patients (Fig. 1A, B, Supplemental Table 2). Other AML-associated somatic variants, such as NPM1, WT1, CEBPA, IDH1, NRAS and DNMT3A were detected for the FLT3-ITD patients, often in a patient-specific manner. For both AML1-ETO patients, the WES analysis revealed a KIT mutation that is associated with poorer prognosis and increased risk of relapse [18, 19].
Fig. 1
Whole exome- and gene fusion analysis between Dx and Re. (A) Oncoplot from WES showing 14 selected somatic mutations across 6 patients (red: n = 2 AML1-ETO; blue: n = 4 FLT3-ITD). We termed patient s232, s292, s2275 and s3432 as “FLT3-ITD”, although it is not an AML initiating lesion, nor an acknowledged WHO2016 AML category, but treatment with the FLT3 inhibitor Midostaurin is distinct from the AML1-ETO patients. Mutations with at least 5 reads on the ALT allele and VAF ≥ 0.05 are depicted as squares and the ones below this threshold are indicated as triangle. Vertical bars depict the number of mutations detected per sample; horizontal bars depict the (relative) frequency of a particular mutation. (B) Gene fusions detected from bulk RNA-seq. (C) Mutations with a VAF ≥ 0.2 at Dx or Re for which the VAF changed significantly. For all bars, p < 0.05, Fisher’s exact test with Benjamini–Hochberg correction. Red: mutations more abundant at Dx. Blue: mutations more abundant at Re
Next, to identify clonal rearrangements that may have led to disease relapse, we screened for somatic mutations with a significantly altered variant allele frequency (VAF) between Dx and Re (VAF ≥ 0.2 and p < 0.05, Fisher’s exact test; methods; Fig. 1C). In the FLT3-ITD group, patient s232, WES and PCR analysis revealed two distinct FLT3-ITD mutations in the Dx sample, one of which one was lost at Re (p = 1.0 × 10–3; Fisher’s exact test based on WES reads; Fig. 1C, Supplemental Table 2). WES also revealed a NPM1 mutation (type A19) at Dx, whose AF was decreased at Re (p = 8.2 × 10–3) and a low-abundant NRAS mutation at Dx (VAF = 0.087) that was undetected at Re (VAF = 0; p = 2.0 × 10–4; Fig. 1C). For patient s2275, WES showed considerably shorter ITDs at Re compared to Dx (p = 4.6 × 10–41), which were confirmed by PCR (Supplemental Table 3). This patient (s2275) had a NUP98-NSD1 fusion at Dx and Re, consistent with the absence of DNMT3A and NPM1 mutations [20, 21]. Finally, this patient displayed copy neutral loss of heterozygosity at the q-arm of chr13 (13q-LOH), which increases the allelic burden of the FLT3-ITD [22]. This aberration existed in a small fraction of Dx cells, but its abundance and that of the FLT3-ITD allele increased drastically at Re (Supplemental Fig. 1, Supplemental Tables 2 and 3). Patient s292 displayed NPM1 and DNMT3A mutations that remained unaltered between DX and Re. Patient s3432 showed a retention of the FLT3-ITD, both in the insertion location and allelic ratio (WES and PCR). Somatic mutations in FAT3 (VAF = 0.238, p = 4.3 × 10–8), ITGB7 (VAF = 0.165, p = 1.32 × 10–6), UBA2 (VAF = 0.117, p = 6.32 × 10–3) and SLC4A3 (VAF = 0.135, p = 6.6 × 10–3) were significantly gained in the Re sample (Fig. 1C and Supplemental Table 2), whereas the last 53 Mb of chr7 was lost on one allele (Supplemental Fig. 1).
One of the AML1-ETO samples (s914) showed two distinct KIT mutations (VAF = 0.325; VAF = 0.138) at Dx, both of which were significantly reduced at Re (p < 4.7 × 10–7). Patient s220 gained a mutation in the FAT3 gene as well as copy number aberrations at chr2 and chr15 at Re (Supplemental Fig. 1).
To summarize, we confirmed the presence of FLT3-ITDs and AML1-ETO in four and two patients respectively. Additional somatic aberrations in AML-associated genes were patient-specific. FLT3-ITDs were altered in two patients. In one patient, one of the two FLT3-ITDs was lost at Re. For patient s232, a NPM1 mutation was detected at Dx (VAF = 0.6), but was decreased at Re (VAF = 0.1, p = 0.008, Fisher’s exact test). We also observed a significant reduction in two distinct KIT mutations in patient s914 between Dx and Re as well as patient-specific copy number changes at Re.
Single cell transcriptomics reveals distinct AML-phenotypes at Dx and ReNext, to better understand the transcriptional phenotypes, their differences and possible mechanisms that led to disease progression, we profiled bone marrow cells obtained at Dx and after Re using single cell transcriptomics. In brief, single CD33+ or CD34+ bone marrow cells were FACS-sorted into 384-well plates following the SORT-seq method [10], we acquired 5,612 single cell profiles, in which 4,129 unique transcripts from 1,678 genes were detected on average (Supplemental Fig. 2A, methods).
After normalization, cells were clustered and visualized using the uniform manifold approximation and projection [23] (UMAP). AML1-ETO vs FLT3-ITD samples are separated by UMAP1 and Dx-Re pairs cluster relatively close together (Fig. 2A, B). Nevertheless, considerable heterogeneity between and within pairs exists (Fig. 2B). Strikingly, Dx-Re cells of FLT3-ITD patient s232 cluster in close proximity suggesting minor phenotypic and molecular alterations, even though this patient lost NPM1 and NRAS mutation at Re. Similarly, the expression changes between Dx and Re cells of patient s914 were minor, despite the significant loss of two KIT mutations. In contrast, Dx cells of patient s3432 are completely separated from Re cells, although one mutation in the FAT3 gene was detected in Re (VAF = 0.238) (Supplemental Table 2). Likewise, the Dx and Re cells of AML1-ETO patient s220 constitute distinct clusters, but only gained mutations in genes that are not associated with AML (Fig. 1C). Both patients did however display large copy number changes between Dx and Re (Supplemental Figs. 1 and 6A). Inferred CNV analysis confirmed that these aberrations were undetected for cells at Dx, but present for almost all cells at Re (Fig. 2C). As expected, these CNVs caused differential gene expression at these loci (Fig. 2D), such as LOXL1 and FAM81A implicated in cancer progression [24].
Fig. 2
Single cell transcriptomics reveals distinct AML-phenotypes. (A) UMAP of the six AML pairs, colored by primary mutation (red: AML1-ETO; blue: FLT3-ITD); (B) UMAP colored by sample; (C) Copy number variation data derived from WES (left) and scRNA-seq (right) data for patient s220. Left: Relapse-specific copy number gain and loss at chr2 and chr15, respectively. Right: cell normalized gene expression signals (iCNV signal) in tiles of 3 Mb show the copy loss and gain at chr2 and chr15, respectively. The plot indicates that virtually all Re cells are affected, compared to none of the Dx cells. Bottom: tumor allele frequency at heterozygous SNPs confirms copy loss at chr2 and gain at chr15. (D) Boxplot of DEGs at the lost and gained segments of chr2 (n = 17 DEGs) and chr15 (n = 24 DEGs), respectively. (E) Heatmap showing the top 20 marker genes per primary mutation
Next, we looked for gene signatures that discriminated AML1-ETO or FLT3-ITD patients. These signatures include well-established AML1-ETO markers, like upregulation of the transcriptional co-repressor RUNX1T1 (aka ETO), the transcription factor POU4F1 [25] and the myeloid differentiation protein MPO [26] (Fig. 2E, top). FLT3-ITD samples on the other hand are characterized by induced expression of VIM, ANXA1, MSI2 and LAPTM5. Other genes tend to be overexpressed only in a subset of the samples: HLA genes are overexpressed in AML1-ETO patient s220, but not in s914. In the FLT3-ITD samples, HOXA5 and HOXB3 genes that are overexpressed in NPM1-mutated AML [27], appear overexpressed in a patient-specific manner (Fig. 2E, bottom). Closer inspection of these and other NPM1-marker genes showed that these genes are indeed significantly higher expressed in the FLT3-ITD samples with an additional NPM1 mutation (NPM1mut) compared to NPM1WT samples (FC > 1.5 and p < 6.0 × 10–15; Supplemental Fig. 2D). Notably, HOX-genes are also highly expressed in FLT3-ITD patient s2275. In these samples, we detected a NUP98-NSD1 fusion gene that is characterized by upregulation of HOXA and HOXB genes [28] (Supplemental Fig. 2D).
In summary, single cell transcriptomics showed distinct clustering of AML1-ETO vs FLT3-ITD patients. Differential analysis confirmed upregulation of well-established marker genes as well as elevated expression of HOX genes in NPM1mut and the NUP98-NSD1 positive FLT3-ITD samples. On a global level, the transcriptional changes between Dx and Re are poorly explained by SNPs and INDELs in AML-associated genes, but rather seem to be associated with large-scale CNVs. To gain a deeper understanding of the mechanisms underlying these changes, we subsequently performed an in-depth analysis of Dx-Re pairs per AML-subtype and in a patient-specific setting.
Dx-Re transcriptomic changes are patient specificGiven this high intra- and inter-patient heterogeneity, we focused on the Dx-Re differences per patient in the remainder of this study. For this, we separated the UMAPs of the FLT3-ITD and AML1-ETO patients (Fig. 3A, B) and computed the differentially expressed genes between the Dx-Re pairs per patient. This analysis reinforced the notion that the differences in transcription between Dx and Re are highly patient-specific (Supplemental Figs. 2B, C, 3C, D).
Fig. 3
Single cell transcriptomics reveals heterogeneity amongst patients. (A) UMAP of the four sample pairs with a FLT3-ITD, colored by sample (red: Dx; blue: Re); (B) UMAP of the two AML1-ETO sample pairs, colored by sample; (C) Heatmap displaying the top 5 marker genes per sample (FLT3-ITD); (D) Heatmap displaying the top 10 marker genes per sample
The FLT3-ITD patients show a modest separation between the Dx and Re samples of patient s232 (Fig. 3A). Cluster analysis revealed two clusters at diagnosis (cluster 1–2) and one at relapse (cluster 3, Supplemental Fig. 3A). Re cells lost expression of members of the AP-1 transcription factor, like FOS, FOSB and ATF3 that were highly expressed in Dx cluster 1 (Supplemental Fig. 3B). Gene ontology (GO) analysis confirmed significant loss of expression for these and other genes involved in AP-1/ATF-2 related transcription at Re (Supplemental Fig. 3C). Furthermore, we evaluated the expression level of genes involved in PI3K/AKT/mTORC pathway, in which mTORC1 controls ribosomal biogenesis and protein translation. We found the targets of mTORC1, like RPS6KB1 and EIF4E, were differentially expressed at Re (Supplemental Fig. 3D), suggesting a pathway shift from AP-1 to mTORC1. Besides, we observed the upregulation of the upstream K/NRAS genes in Re, which may be markers for diagnosis/prognosis and treatment targets.
The UMAP for patient s292 showed 3 distinct clusters (Supplemental Fig. 4A). DEG between Dx clusters 1 and 2 revealed IDH1, an enzyme in the TCA cycle, and RAB31 involved in membrane fusion and exocytosis in clusters 1, whereas MPO and PROM1, markers for GMP cells, are differentially expressed in cluster2 (Supplemental Fig. 4B, C). Cells in cluster 3 originate from the Re sample and overexpressed genes like DDIT4 [29], PIM3 [30] and CD74 [31] were previously associated with poor prognosis (Supplemental Fig. 4B). GO analysis indicated regulation of cell death and apoptotic process terms in cluster 3 (Supplemental Fig. 4C).
The copy-neutral LOH at chr13q could be detected from the alternate allele frequency in the WES and scRNA-seq data of patient s2275 (Supplemental Fig. 5A). However, no genes located on chr13q were differentially expressed at Re compared to Dx, with the exception of ELF1 (twofold downregulated at Re). Single cell expression analysis revealed 5 clusters. Cluster 1 mainly originated from Dx cells, whereas cluster 5 almost entirely consisted of Re cells. Clusters 2–4 however were a mixture between Dx and Re cells (Supplemental Fig. 5B, C). DEGs revealed few differences between cluster 1 and 5, such as RNU4ATAC and RYBP involved in RNA biosynthesis and metabolism that are differential expressed in cluster 1 (Dx) (Supplemental Fig. 5C, D), whereas ITM2A and CLEC12A for leukocyte activation and LDHA for ribonucleotide metabolism are differentially expressed in cluster 5 (Re) (Supplemental Fig. 5C, D). The minor differences between Dx and Re is consistent with the fact that AML-associated mutations, such as FLT3-ITD, WT1, CEBPA and NUP98-NSD1 were present at Dx and retained at Re (Fig. 1A, B, Supplemental Table 2).
Fig. 4
Pathway switch between AP-1 and RAS signaling in high risk FLT3-ITD (s3432). (A) UMAP of Dx and Re cells for FLT3-ITD patient s3432 colored by timepoint (top) or cell cluster (bottom). (B) Heatmap displaying the top 10 cluster marker genes. Color represents row normalized expression values. (C) Overrepresented GO terms (category: biological pathway) in cluster 1 (Dx) and 3 (Re). P-values: hypergeometric test (BH-corrected). (D) The expression of genes related to AP-1 transcription factor network and RAS signaling pathway in each timepoint. (E) Calculation of LSC17 score for each cluster, and p-value was calculated using Student’s t-test. * p < 0.05, ** p < 0.01, *** p < 0.001
The Dx and Re cells of patient s3432 formed distinct clusters that are highly separated from each other and the other FLT3-ITD patients (Fig. 3A). Cluster analysis detected four groups of cells that largely separated Dx (cluster 1) from Re cells (cluster 2–4; Fig. 4A). Cluster 1 had a characteristic gene signature of transcription factors involved in proliferation and cell growth (e.g., JUN, FOS, FOSB, EGR1, SOX4 and KLF6) that were significantly downregulated in the Re clusters (Fig. 4B, C). The Re-specific clusters 3–4 upregulated genes involved in the RAS/mTORC pathway, such as ANKRD28 and PIK3R1, whereas cluster 2 is hallmarked by cell cycle related genes, such as TOP2A and MKI67. Pathway enrichment analysis confirmed the overrepresentation of AP-1/ATF2 transcription factors in cluster 1 (Dx) and additionally revealed upregulation of genes involved in mTOR signaling, like RICTOR, PIK3R1 and HIF1A in cluster 3 (Re; Fig. 4C, D). This suggests a pathway switch from AP-1 in the diagnosis cells towards mTOR in the relapse cells. We further observed that KRAS and NRAS, genes upstream of mTORC, were also overexpressed in the Re sample (Supplemental Fig. 6C). Interestingly, cluster 4 in relapse is characterized by elevated exocytosis (Supplemental Fig. 6D) and increased expression of genes related to Tim-3-galectin-9 Secretory Pathway (e.g. ADGRL1, HAVCR2 and LGALS9) that protect AML cells against from the host immune system in an mTOR dependent manner [32] (Supplemental Fig. 6E), in particular from NK- and T-cell action. Finally, the leukemia stem cell (LSC) score, a 17-gene signature (LSC17) that correlates with aggressiveness of the leukemia and a poor outcome [16] was significantly higher in the Re clusters 3 and 4 compared to the Dx cluster 1 (Fig. 4E). For the other FLT3-ITD patients, none of the clusters had an elevated LSC17 score (data not shown).
Fig. 5
Putative LSCs detected in AML1-ETO pair (s914). (A) UMAP of Dx and Re cells for AML1-ETO patient s914, colored by timepoint (top) and cell cluster (bottom). Cells in cluster 6 express ambiguous marker genes, and may be doublets or contaminated by ambient RNA and were discarded (see also Supplemental Fig. 6). (B) Heatmap depicting the top 7 cluster markers. Color represents row normalized expression values. (C) Pseudo-time trajectory colored by timepoint (top) or cell cluster (bottom). (D) Heatmap showing representative genes per cluster. (E) LSC17 score per cluster. * p < 0.05, ** p < 0.01, *** p < 0.001, Student’s t-test. (F) Barplots depicting the relative cell abundance per cell cycle phase (inferred from marker gene expression) for each cell cluster. Arrow: cells in cluster 2 and 3 predominantly reside in the G1 phase
Leukemic stem cell-like cells in AML1-ETOHigher MPO, a marker for granulocyte/monocyte progenitors (GMPs) expression [26] within both the AML1-ETO patients (Fig. 2E) implies that most cells are arrested at a “GMP-like” stage. The Dx and Re cells of AML1-ETO patient s220 were more separated compared to those of patient s914 (Fig. 3B, Supplemental Fig. 2C). 41 of the DEGs are transcribed at the amplified or lost loci and their altered transcription may in turn deregulate other genes.
Analysis on Dx-Re showed that the number of DEGs shared between these two AML1-ETO patients is minimal as for the FLT3-ITDs (Supplemental Fig. 2C). Therefore, we performed an in-depth analysis on the transcriptional dynamics between Dx and Re separately for these two patients. Focusing on patient s914 first, the synergic oncogenes (PIM1 and MYC [33]) responsible for tumorigenesis were co-differentially expressed at Re compared to Dx. Cluster analysis revealed five groups of cells (Fig. 5A, B) and a small cluster of scattered cells that expressed signatures of progenitors (CD34), erythrocytes (HBB), monocytes (LYZ), B-cells (MSA41) and cell cycle related genes (TOP2A, MKI67) (Supplemental Fig. 7) likely resulting from ambient RNA or cell doublets and hence were discarded in subsequent analyses.
Cluster 1 mainly consist of Dx cells and differentially expressed genes for differentiation and resistance to apoptosis, like AREG [34]. Interestingly, cells in cluster 2 express CD34 as well as genes involved in cell migration (ANXA1 [35], ANXA2 [36], VIM [37] and EMP1 [38]) but lacked the expression of MPO (Fig. 5B, D). To investigate whether and from which Dx cluster these potential Re LSCs originate, we aligned cells in pseudo-time based on the gradient of transcriptional differences using Monocle3. This trajectory analysis suggested a continuous transition between the Dx and Re sample (Fig. 5C). Cells in cluster 2 and 3 differentially expressed genes for hematopoietic stem cell maintenance (GDF11 [39], GATA2 [40]) and differentiation (GAS7 [41], CAMK1D [42]) markers as well as CD34 (Fig. 5B, D), indicating cluster 2 and 3 are the putative starting points of this trajectory. Besides, cluster 2 and 3 overexpressed genes CXCR4 [43] and CXCL8 [44] for tumor microenvironment (Fig. 5B, D). In line with those findings, we calculated the LSC17- and cell cycle scores for all clusters. We observed that cells in Dx cluster 3 have the highest LSC17 score followed by Re cluster 2 (Fig. 5E). Moreover, cells from cluster 2 and 3 mainly reside in the G1 phase of the cell cycle (Fig. 5F). Interestingly, the trajectory suggest that these cells differentiate into a population of cells that display DUSP6 and AP-1 related genes like JUN and FOS in the Re-specific clusters 3 and 4 (Fig. 5D).
UMAP shows that s220 cells separate according to Dx and Re which partitioned into 9 clusters (Fig. 6A). Clusters 1–4 contained Dx cells that were enriched for CXCL8 and CXCR4, genes associated with the interaction between leukemia blasts and stromal cells [43, 44]. Clusters 5–9 exclusively contained Re cells and were marked by expression of LOXL1 and FAM81A (Fig. 6B). Cell cycle-related genes (MCM6, TOP2A, MKI67) were highly expressed in cluster 1 and 9. Cluster 4 (Dx) and 5 (Re) are in close proximity to each other and share marker genes, such as CAMK1D, GAS7, ANXA1/2, VIM and CD34 (Fig. 6B, Supplemental Fig. 8A) suggesting that they are LSCs.
Fig. 6
Putative LSCs detected in AML1-ETO pair (s220). (A) UMAP of Dx and Re cells for AML1-ETO patient s220, colored by timepoint (top) and cell cluster (bottom). (B) Heatmap depicting the top 5 marker genes per cluster. Color represents row normalized expression values. (C) LSC17 scores per cluster. * p < 0.05, ** p < 0.01, *** p < 0.001, Student’s t-test. (D) top: Barplots depicting the relative cell abundance per cell cycle phase (inferred from marker gene expression) for each cell cluster. Arrow: cells in cluster 4 and 5 predominantly reside in the G1 phase. Bottom: UMAP colored by cell cycle phase. (E) Pseudo-time trajectory colored by cell cluster (F) Heatmap depicting representative marker genes per cluster/inferred timepoint
Alternative “branching” from Re and Dx LSC-like cells in AML1-ETOGiven the high similarities between clusters 4 and 5 and their elevated CD34 expression, we hypothesized that these clusters might be enriched in LSCs. Analysis showed that these clusters indeed have the highest LSC17-score and contain cells that reside predominantly in the G1 cell cycle phase (Fig. 6C, D). To better understand the transcriptional dynamics of cell populations originating from these LSCs, we applied pseudo-time gene expression analysis (Fig. 6E). This analysis reveals a trajectory starting from the presumed LSCs cluster 4 and 5 towards more differentiated cells that predominantly reside in the S-phase of the cell cycle and exhibit elevated expression of genes like TOP2A and MKI67 (Fig. 6D-F). For the Dx branch, genes involved in self-renewal that impede differentiation (GAS7 and CAMK1D) or are associated with cell migration (TPPP3, VIM, ANXA1/2) had elevated expression in cluster 4. We hypothesized that all other clusters of cells originate from this presumed Dx LSC population. Indeed, we observed a downregulation of these markers when cells are traced along the trajectory from cluster 4 to cluster 1 which is consistent with their differentiation into more mature myeloid cells. Furthermore, DUSP1 and DUSP6, genes required for cell differentiation and proliferation were upregulated as cells ‘moving away’ from cluster 4 along the Dx branch (Fig. 6F). In the Re branch, TPPP3, VIM, ANXA1/2, GAS7 and CAMK1D were upregulated in cluster 5 to a similar extent as in cluster 4. Compared to the more gradual downregulation in the Dx branch, these markers were largely lost when cells “branched” from cluster 5 to cluster 6 (Fig.
留言 (0)